|
|
 |
| Little spikemoss. Image by Dave Powell, USDA Forest Service, Bugwood.org. |
Introductory
SPECIES: Selaginella densa
AUTHORSHIP AND CITATION :
Crane, M. F. 1990. Selaginella densa. In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station,
Fire Sciences Laboratory (Producer). Available:
www.fs.usda.gov/database/feis//plants/fern/selden/all.html [].
Revisions:
On 5 April 2018, the common name of this species was changed in FEIS
from: little spikemoss
to: lesser spikemoss. Images were also added.
ABBREVIATION :
SELDEN
SYNONYMS :
Selaginella rubestris var. densa
Selaginella scopulorum
Selaginella engelmannii
Selaginella engelmannii var. scopulorum
Selaginella engelmannii var. standleyi
Selaginella standleyi
NRCS PLANT CODE :
SEDE2
SEDED
SEDES
SEDES2
COMMON NAMES :
lesser spikemoss
little clubmoss
little spikemoss
prairie spikemoss
Rocky Mountain selaginella
spikemoss
TAXONOMY :
The scientific name of lesser spikemoss is Selaginella densa Rydb.
(Selaginellaceae). There are three varieties of lesser spikemoss [31]:
Selaginella densa var. densa, lesser spikemoss
Selaginella densa var. scopulorum (Maxon) Tryon, Rocky Mountain spikemoss
Selaginella densa var. standleyi (Maxon) Tryon, Standley's spikemoss
LIFE FORM :
Fern Ally
FEDERAL LEGAL STATUS :
No special status
OTHER STATUS :
NO-ENTRY
DISTRIBUTION AND OCCURRENCE
SPECIES: Selaginella densa
GENERAL DISTRIBUTION :
The range of lesser spikemoss extends south from the Alaska panhandle to
northwest California and east into Manitoba and the Dakotas. It is
common on the Great Plains and in the Rocky Mountains it grows as far
south as eastern Utah and Arizona, New Mexico and Trans-Pecos Texas
[13,27].
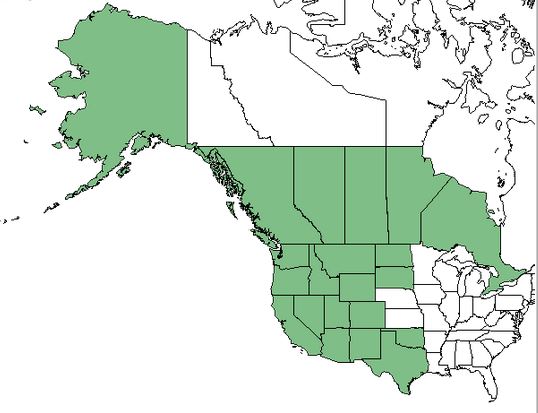 |
| Distribution of lesser spikemoss. Map courtesy of USDA, NRCS. 2018. The PLANTS Database.
National Plant Data Team, Greensboro, NC [56] [2018, April 4]. |
The distributions of the varieties of lesser spikemoss are as
follows [13,26,27,42]:
Selaginella densa. var. densa is the characteristic variety found on the
northern Great Plains. It may occasionally extend into western Montana, southwest
Alberta and southeast British Columbia. Further south it is found in
the mountains of Colorado and the LaSal Mountains of Utah.
Selaginella densa var. scopulorum is the common variety in the mountains of the
Northern Rocky Mountain Region through the LaSal and Uinta mountains of
eastern Utah to west Texas. It is also found from British Columbia to
northern California. It is less common in the mountains of Colorado.
Selaginella densa var. standleyi is uncommon and found only above timberline in the
northern Rocky Mountains.
ECOSYSTEMS :
FRES10 White - red - jack pine
FRES20 Douglas-fir
FRES21 Ponderosa pine
FRES23 Fir - spruce
FRES26 Lodgepole pine
FRES29 Sagebrush
FRES30 Desert shrub
FRES34 Chaparral - mountain shrub
FRES35 Pinyon - juniper
FRES36 Mountain grasslands
FRES38 Plains grasslands
FRES39 Prairie
FRES40 Desert grasslands
FRES44 Alpine
STATES :
AZ CA CO KS MT NE NM ND OK OR
SD UT WA WY AB BC MB SK
BLM PHYSIOGRAPHIC REGIONS :
1 Northern Pacific Border
2 Cascade Mountains
4 Sierra Mountains
5 Columbia Plateau
8 Northern Rocky Mountains
9 Middle Rocky Mountains
10 Wyoming Basin
11 Southern Rocky Mountains
12 Colorado Plateau
13 Rocky Mountain Piedmont
14 Great Plains
15 Black Hills Uplift
16 Upper Missouri Basin and Broken Lands
KUCHLER PLANT ASSOCIATIONS :
K007 Red fir forest
K008 Lodgepole pine - subalpine forest
K011 Western ponderosa forest
K012 Douglas-fir forest
K015 Western spruce - fir forest
K016 Eastern ponderosa forest
K017 Black Hills pine forest
K018 Pine - Douglas-fir forest
K019 Arizona pine forest
K020 Spruce - fir - Douglas-fir forest
K021 Southwestern spruce - fir forest
K023 Juniper - pinyon woodland
K037 Mountain mahogany - oak scrub
K038 Great Basin sagebrush
K039 Blackbrush
K040 Saltbush - greasewood
K051 Wheatgrass - bluegrass
K052 Alpine meadows and barren
K055 Sagebrush steppe
K056 Wheatgrass - needlegrass shrubsteppe
K057 Galleta - three-awn shrubsteppe
K063 Foothills prairie
K064 Grama - needlegrass - wheatgrass
K065 Grama - buffalograss
K066 Wheatgrass - needlegrass
K067 Wheatgrass - bluestem - needlegrass
K070 Sandsage - bluestem prairie
K095 Great Lakes pine forest
SAF COVER TYPES :
1 Jack pine
205 Mountain hemlock
206 Engelmann spruce - subalpine fir
207 Red fir
208 Whitebark pine
209 Bristlecone pine
210 Interior Douglas-fir
218 Lodgepole pine
219 Limber pine
220 Rocky Mountain juniper
237 Interior ponderosa pine
238 Western juniper
239 Pinyon - juniper
SRM (RANGELAND) COVER TYPES :
NO-ENTRY
HABITAT TYPES AND PLANT COMMUNITIES :
In the Great Plains grasslands of southern Canada and the northern
United States, lesser spikemoss can be a dominant in three of the five
major grass associations: the short-grass prairie, mixed prairie and
fescue (Festuca scabrella) prairie [7,9]. At higher elevations in
Glacier Park, it codominates with Idaho fescue (F. idahoensis) in a
climax grassland association [48]. In the Custer National Forest of
Montana, North and South Dakota and the Theodore Roosevelt National Park
of North Dakota, lesser spikemoss is rare outside of the needle-and-thread grass
(Hesperostipa comata)/threadleaf sedge (Carex filifolia) habitat
type where it is an important species [24,25]. Published classification
schemes listing lesser spikemoss as an indicator species or a dominant
part of vegetation in plant associations (pas) are presented below:
Area Classification Authority
Region 2: CO,NE,KS, general veg. pas Johnston 1987
SD,WY
MANAGEMENT CONSIDERATIONS
SPECIES: Selaginella densa
IMPORTANCE TO LIVESTOCK AND WILDLIFE :
Lesser spikemoss is a minor part of elk diets but is not a valuable elk
browse [36]. During a Montana study domestic animals were never
observed grazing on lesser spikemoss [57]. Bighorn sheep may eat some
spikemoss (Selaginella spp.) in New Mexico [40].
PALATABILITY :
The degree of use shown by livestock and wildlife species for
lesser spikemoss (var. densa) in several western states is rated as
follows [16]:
UT WY MT ND
Cattle poor poor poor poor
Sheep poor poor poor poor
Horses poor poor poor poor
Elk fair poor poor ----
Mule deer fair poor poor fair
White-tailed deer ---- poor ---- fair
Antelope fair poor poor fair
Upland game birds fair fair ---- poor
Waterfowl fair ---- ---- ----
Small nongame birds fair ---- ---- ----
Small Mammals fair fair ---- ----
NUTRITIONAL VALUE :
The energy and protein value of lesser spikemoss forage is poor [16].
Aboveground parts of lesser spikemoss have a high ash content (19.7%).
Most of this is silica which comprises about 16.5 percent of plant dry
weight. Other results of this analysis are given below [59]:
Percent of dry weight of aboveground portions of plant
Calcium 0.20
Potassium 0.30
Iron 0.038
Zinc 0.023
Sodium 0.29
Phosphorus 0.11
Protein 8.50
COVER VALUE :
Lesser spikemoss provides poor cover for small animals [16].
VALUE FOR REHABILITATION OF DISTURBED SITES :
Lesser spikemoss has poor short-term or long-term revegetation potential
[16]. It does grow well on gentle and moderate slopes and it has low to
moderate erosion control potential. However, its potential biomass
production is low and establishment requirements may be exacting [16].
OTHER USES AND VALUES :
While lesser spikemoss does not provide forage, it does protect the soil
from erosion by wind or water and reduces the effects of tramping [7].
With care, lesser spikemoss can be transplanted into gardens where it is
an attractive ground cover that grows on rocks and in other dry
situations [34].
OTHER MANAGEMENT CONSIDERATIONS :
The role of lesser spikemoss in mixed prairie is not well understood,
although its influence on other vegetation and the habitat has been
thought to be slight because its requirements for water and nutrients
are low [9,10,60]. During light showers, its dense mats and extensive,
shallow roots may intercept and soak up all the available water [55,57].
During high intensity storms, the mats of lesser spikemoss act to
increase infiltration and decrease runoff and surface erosion [57]. Its
shallow root system also limits spikemoss growth to periods of the year
when moisture is available at shallow depths [59].
Grazing effects: The effect of grazing on lesser spikemoss seems to be
confounded by drought. In Canadian and Montana mixed prairie, lesser
spikemoss is able to increase during periods of drought [10,18,58].
Also, lesser spikemoss increases when protected from grazing and
decreases with grazing due to trampling of grazing animals [10]. Lesser
spikemoss is relatively weakly rooted and easily damaged by trampling
[37]. Decreases in response to grazing may also be explained by the
exposure and drying of the surface soil when cover is removed by grazing
[18]. In the short-grass prairie of southeastern Alberta and
southwestern Saskatchewan, grasses can easily establish in areas with
high cover of lesser spikemoss during years with above normal
precipitation, but they are rarely able to establish during dry years
[7].
A 4-year Montana study done during a period of normal rainfall compared
ungrazed mixed prairie with mixed prairie moderately grazed by sheep.
Cover of lesser spikemoss decreased from 21 to 17.6 percent on the
ungrazed site but it decreased from 15.1 to 7.1 percent on the grazed
area [58]. The strong decrease on grazed prairie was attributed to
trampling, competition, and manure [57].
A 7-year study of three levels of grazing intensity on the short-grass
prairie in southeastern Alberta and southwestern Saskatchewan was done
during dry and drought years [9]. Lesser spikemoss increased under all
levels of grazing, with the greatest increase in the most lightly grazed
area. On ungrazed land lesser spikemoss increased in basal area from 11
percent to 16 percent. Results of a second 10-year study showed the
basal area of lesser spikemoss increased by 37 percent on ungrazed and 27
percent on grazed land, while grass cover generally decreased. Looking
at total vegetation cover the authors concluded that "climate rather
than moderate grazing use was the principal factor affecting plant
cover" between 1928 and 1939. During a third 5-year study, lesser
spikemoss increased more under a medium level of grazing than under
severe grazing intensity [9]. Lesser spikemoss increases more under
rotational grazing than under continuous grazing [9,65].
A 3-year North Dakota mixed prairie study compared exclosures
established 38 to 41 years previously with cattle-grazed range. Lesser
spikemoss cover did not vary significantly despite major differences in
species composition of grasses and silver sagebrush (Artemisia cana)
[5,6]. Site differences were more important than grazing pressure [6].
Ross and Hunter [50] list spikemoss as increasing under grazing pressure
in Montana. However, lesser spikemoss increases very slowly when taller,
competing vegetation is partly or wholly removed [52].
Treatment with fertilizer: The primary effect of manure or fertilizer
applications is to increase the competitiveness of taller grasses, thus
decreasing lesser spikemoss [18]. However, in the northern plains
response depends upon precipitation [54]. There is no evidence that
nitrogen application alone kills lesser spikemoss [52]. When fertilizers
containing nitrogen and phosphorus were added to grassland plots in each
of 2 successive years, the cover of live lesser spikemoss decreased
significantly. It was not determined whether this was due to the
nutrients themselves or because of increased competition from other
vegetation [57]. Manuring alone increased all plant cover the first
season of a Montana study, but lesser spikemoss was subordinated to the
more vigorous native grasses and decreased the second season [18].
After more than 30 years with normal weather, lesser spikemoss was still
reduced in plots treated by annual applications of manure for 1 to 11
years [19]. Mulching, with or without added fertilizer, can nearly
eliminate lesser spikemoss [66].
Mechanical treatment: Mechanical treatments reduced lesser spikemoss
cover between 25 and 70 percent in northern Montana mixed prairie [52].
The effects of various combinations of manure treatment, disking, and
harrowing over a 10-year period along with seeding of crested wheatgrass
(Agropyron cristatum) and yellow sweetclover (Melilotus officinalis)
have been studied [18,19]. Most treatments resulted in long-term
decreases in lesser spikemoss, and combination treatments seemed most
successful. When the treated area was resurveyed over 30 years later,
lesser spikemoss cover was still so decreased that researchers concluded
that lesser spikemoss does not easily reestablish on northern mixed
prairie following mechanical treatment [19]. Lesser spikemoss cover and
the cover of either other vegetation or litter were significantly and
negatively correlated [19].
Chemical control: The most effective chemicals for control of lesser
spikemoss while increasing the yield of desirable grasses are AMS
(ammate) and atrazine [59]. Spring treatment is most effective.
Monuron, paraquat, and bromacil control lesser spikemoss but also reduce
the yield of or kill other vegetation [59]. Further range tests
indicate that ammate, atrazine, and monuron are the best treatments.
Their effectiveness is increased with the use of a surfactant
[52].
BOTANICAL AND ECOLOGICAL CHARACTERISTICS
SPECIES: Selaginella densa
GENERAL BOTANICAL CHARACTERISTICS :
Description: Lesser spikemoss is an evergreen, nonflowering herb that
forms dense, cushionlike mats which are seldom more than 1 inch (2.5 cm)
in height [10,57]. The short, leafy, compactly branched stems lie along
the soil surface and may be 4 inches (1 dm) across [27]. The simple,
awn-tipped leaves are very small, up to about 0.1 inch (2.5 mm) long and
0.02 inch (0.5 mm) wide, and arranged in a relatively dense spiral along
the stem. There is usually a single, unbranched vein. The leaves
underneath the stem are longer than leaves on top along the same part of
the stem [42]. Lesser spikemoss has true roots which are very fine
(0.008 inch or 0.2 mm in diameter) and minutely branched. They form a
tangled mass and may comprise 86 percent of the plant dry matter [59].
Most roots occur within 0.78 to 2.0 inches (2-5 cm) of the soil surface
[12].
Drought Resistance: Lesser spikemoss is very resistant to drought if the
roots are not disturbed. In the laboratory, clumps allowed to desiccate
for 6 and 33 months were then revived simply by watering and later
planting [55,61]. The clump that was revived after 33 months developed
reproductive cones within 3 months [61]. When all soil was removed from
roots, no plants survived after 5 weeks of air drying [59]. Shoots that
are dormant during drought have their leaves closely appressed to the
stem and are a dull grayish-green instead of the ash-gray of dead shoots
[61]. Young plants are equally drought resistant. On a site near
Saskatoon, Saskatchewan, Webster and Steeves [61] found over 50
locations where sporelings had survived periods of drought.
RAUNKIAER LIFE FORM :
Chamaephyte
REGENERATION PROCESSES :
The spikemoss life cycle differs from that of flowering plants. At
branch ends, four-angled "cones" or strobili are formed. Each has two
types of spore-bearing structures, microsporangia and megasporangia, in
the leaf axils, with megasporangia located below the microsporangia
[61]. Microspores are released passively to fall through the strobili
to the ground [32]. At maturity they will release microscopic,
flagellated sperm cells. The sperm require water to swim to the egg cell
which develops from cells of the megasporangia. Where fertilization
occurs is not known [52]. Tissue surrounding the fertilized egg may
carry on photosynthesis as well as provide stored food for the
developing embryo. This entire structure may be shed or held in the
leaf axils of the strobili while the embryo develops. The necessity of
water for sexual reproduction restricts the habitat of spikemosses [15],
and young plants are not often reported [18,52].
As lesser spikemoss clumps grow the centers die, leaving a ring of live
material surrounding a dead core [59]. Under range conditions lateral
growth was observed to be less than 0.4 inch (1 cm) a year, making
vegetative spreading a slow process [18,59]. During another field study
growth of less than 0.2 inch (5 mm) in 5 seasons was observed [52].
Fragmentation might be another means of vegetative reproduction and
dispersal [18], although stems of lesser spikemoss do not have a natural
tendency to fragment when dry [55].
SITE CHARACTERISTICS :
Lesser spikemoss is most often found on dry shallow soils that are
gravelly or rocky or have gravelly or stony subsoils [57]. It is most
common in grasslands, alpine tundra, and high subalpine communities
which occupy dry, snow-free ridges. Lesser spikemoss is an extremely
stress-tolerant species and can survive in cold alpine regions with a
short growing season, drought, and summer frosts [62]. Lesser spikemoss
is not common in forests but can be found in Douglas-fir (Pseudotsuga
menziesii) with ponderosa pine (Pinus ponderosa) or lodgepole pine (P.
contorta), Engelmann spruce (Picea engelmannii) and subalpine fir (Abies
lasiocarpa), or aspen (Populus tremuloides) communities [33,44]. Lesser
spikemoss also grows in pinyon (Pinus edulis)-juniper (Juniperus spp.),
sagebrush (Artemisia spp.), and krummholz communities [63]. In Jackson
Hole, Wyoming the big sagebrush (A. tridentata) community has abundant
lesser spikemoss [47]. In northern Montana aspen grove and grassland
communities, it is found primarily with Hood's phlox (Phlox hoodii) on
stony moraine summits, outwash fans and coarse gravels [38]. The
abundance of lesser spikemoss in the aspen grove and grassland community
decreases to the north in Saskatchewan [11].
Grassland Sites: Lesser spikemoss is most abundant in short-grass
prairie. Its basal area may be equal to that of all other herbs and
shrubs [7,9]. In mixed prairie, lesser spikemoss is slightly less
abundant, occurring with an overall frequency of 74 to 98 percent and
cover up to 25 percent [10]. It has much less cover and is not evenly
distributed in the fescue prairie [7,11]. In the northern Red River
Valley, it dominates with needle-and-thread grass [45]. On sites with
standing surface water, lesser spikemoss will be unimportant [64].
However, in Montana, areas where it grows are closer to permanent water
than areas where it does not grow [57]. In North Dakota, lesser
spikemoss is common on a variety of grassland habitat types except those
with extra moisture [64].
Alpine and subalpine sites: In the Washington and British Columbia
Cascades, lesser spikemoss is common in herb field, the Bellard
alpinesedge (Kobresia myosuroides), and purple reedgrass (Calamagrostis
purpurascens) communities on sites that are essentially snow-free during
the winter. On such sites it is exposed to high wind, low temperatures,
and frequent frosts all winter [20]. On alpine sites in Montana, lesser
spikemoss does well where Hooker mountainavens (Dryas octopetala) is
dominant [2]. In alpine areas in Utah, it grows in cushion plant
communities with alpine avens (Geum rossii) and on dry meadows and
alpine turf [37]. On the Colorado Front Range, lesser spikemoss is part
of several forest communities, but its greatest abundance is at higher
elevations in Bellard alpinesedge meadows or under limber pine (Pinus
flexilis) on exposed rocky sites [33,44]. It is well adapted to these
and other alpine cushion-plant communities [62]. In contrast, near
Schoolroom Glacier in the Teton Range of Wyoming, it is found in a
meadow community with kentrophyta milkvetch (Astragalus kentrophyta)
[53]. In Alberta it grows under limber pine on the rocky, dry slopes
and summits of exposed ridges [41]. In northern California lesser
spikemoss (var. scopulorum) grows on open rocky sites in red fir (Abies
magnifica) and lodgepole pine forests of Siskiyou County [42].
Parent material: Lesser spikemoss grows in the drier interior of British
Columbia except for a disjunct population on dry, well-aerated and warm
limestone soils on the Queen Charlotte Islands [49]. In Montana, lesser
spikemoss does well on sites with thin, rocky, and unstable soils derived
from calcareous parent material [2]. In the Bighorn Mountains of
Wyoming, it is present on granitic soils but almost absent on soils
derived from sedimentary rock [57].
Soil texture: Lesser spikemoss is common on medium texture soils and
sandy loams. It is less common on sandy soils and rare on clay
[10,52,57]. In grasslands dominated by bluebunch wheatgrass
(Pseudoroegneria spicata) lesser spikemoss is found in areas with
shallower, rockier, and less sandy soils than areas without lesser
spikemoss [57]. Optimum soil depth for lesser spikemoss (var. densa) is
between 10 and 20 inches (25.4-50.8 cm) [16]. Growth of lesser spikemoss
(var. densa) on various soils in several western states is ranked as
follows [16]:
UT WY MT ND
Soil texture
Gravel good fair fair fair
Sand good poor fair fair
Sandy loam good good fair fair
Loam good good good good
Clay loam good good good good
clay fair fair fair good
dense clay poor poor poor poor
Organic soils fair poor poor ----
Acidic soils fair poor poor ----
Saline soils fair poor poor poor
Sodic soils ---- poor poor fair
Sodic-saline soils poor poor poor poor
Elevation: Elevational ranges in some western regions are
[16,26,30,42,63]:
Minimum Maximum
feet meters feet meters
Arizona 11,470 3,496 11,470 3,496
California
var. scopulorum 5,000 1,524 7,000 2,134
Colorado
var. densa 4,000 1,219 12,000 3,658
var. scopulorum 8,000 2,438 12,500 3,810
var. standleyi 9,500 2,896 13,000 3,962
Montana
var. scopulorum 6,300 1,920 6,300 1,920
Utah 8,858 2,700 14,107 4,300
Wyoming
var. densa 6,000 1,829 10,800 3,292
var. scopulorum 5,600 1,707 10,800 3,292
var. standleyi 5,600 1,707 10,800 3,292
SUCCESSIONAL STATUS :
In general, spikemosses (Selaginella spp.) are unable to compete with
other plants in relatively moist habitats [55]. They also are not
important pioneers. Instead spikemosses are found on open, xeric sites
supporting particular plant communities but shift locally within those
communities. On such open sites their ability to withstand desiccation
allows them to survive and grow well [55]. On alpine sites of Montana's
Beartooth Plateau, lesser spikemoss pioneers on the gravel mulch left by
rodent disturbance in grassy meadows [29]. Lesser spikemoss (var.
scopulorum) is found in alpine areas on unstable soils disturbed by
frost and abraded by wind [29].
SEASONAL DEVELOPMENT :
In North Dakota spore production begins in June and finishes in July
[16]. In Canadian mixed prairie, vegetative growth begins in early
April, strobili first appear late in April, and spores mature late in
May [10]. In the Colorado alpine, leaves of lesser spikemoss remain
green all winter when protected from wind but turn brown and erode where
exposed [3]. The cones (strobili) change from green to brown in late
May or early June when they are released from snow [3].
FIRE ECOLOGY
SPECIES: Selaginella densa
FIRE ECOLOGY OR ADAPTATIONS :
Lesser spikemoss would appear to have lesser defense against fire.
However, it has been observed to survive on dry rocky hillsides where
other vegetation and litter were sparse. On nearby moist areas where
prefire vegetation was more dense, it was completely removed [51].
FIRE REGIMES :
Find fire regime information for the plant communities in which this
species may occur by entering the species name in the FEIS home page under
"Find Fire Regimes".
POSTFIRE REGENERATION STRATEGY :
secondary colonizer; off-site spores carried to site after year two
FIRE EFFECTS
SPECIES: Selaginella densa
IMMEDIATE FIRE EFFECT ON PLANT :
Lesser spikemoss is killed by fire except where it is protected by site
conditions [51].
DISCUSSION AND QUALIFICATION OF FIRE EFFECT :
NO-ENTRY
PLANT RESPONSE TO FIRE :
Generally, spikemosses are lacking following recent fire or other major
disturbance [55]. In a study of paired stands in mixed grass prairie in
Manitoba, lesser spikemoss cover was 14.9 percent in a control plot, 6.2
percent following a single fire and 2.3 percent following two fires
[67]. In paired stands in western North Dakota, the frequency of
lesser spikemoss was much less following fires that burned from 3 months
to 4 years before sampling [17].
DISCUSSION AND QUALIFICATION OF PLANT RESPONSE :
NO-ENTRY
FIRE MANAGEMENT CONSIDERATIONS :
Prescribed fire can be used to reduce the cover of lesser spikemoss. Any
method of reducing lesser spikemoss would be most successful if done
during a cycle of wet years on the mixed prairie, as drought would
severely set back the recovery of desirable vegetation [1].
Selaginella densa: References
1. Bailey, Arthur W. 1978. Effects of fire on the mixed prairie vegetation. In: Proceedings: Prairie prescribed burning symposium and workshop; 1978 April 25-28; Jamestown, ND. [Place of publication unknown]: [Publisher unknown]: [5 pages]. On file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. [3598]
2. Bamberg, Samuel A.; Major, Jack. 1968. Ecology of the vegetation and soils associated with calcareous parent materials in three alpine regions of Montana. Ecological Monographs. 38(2): 127-167. [12554]
3. Bell, Katherine L. 1974. Autumn, winter and spring phenology of some Colorado alpine plants. The American Midland Naturalist. 91(2): 460-464. [233]
4. Bernard, Stephen R.; Brown, Kenneth F. 1977. Distribution of mammals, reptiles, and amphibians by BLM physiographic regions and A.W. Kuchler's associations for the eleven western states. Tech. Note 301. Denver, CO: U.S. Department of the Interior, Bureau of Land Management. 169 p. [434]
5. Brand, M. D.; Goetz, H. 1978. Secondary succession of a mixed grass community in southwestern North Dakota. Annual Proceedings of the North Dakota Academy of Science. 32(2): 67-78. [7512]
6. Brand, Michael D.; Goetz, Harold. 1986. Vegetation of exclosures in southwestern North Dakota. Journal of Range Management. 39(5): 434-437. [507]
7. Campbell, J. B.; Lodge, R. W.; Johnston, A.; Smoliak, S. 1962. Range management of grasslands and adjacent parklands in the prairie provinces. Publ. 1133. Ottawa, ON: Canada Department of Agriculture, Research Branch. 32 p. [595]
8. Carroll, S. B.; Bliss, L. C. 1982. Jack pine - lichen woodland on sandy soils in northern Saskatchewan and northeastern Alberta. Canadian Journal of Botany. 60: 2270-2282. [7283]
9. Clarke, S. E.; Tisdale, E. W.; Skoglund, N. A. 1943. The effects of climate and grazing practices on short-grass prairie vegetation in southern Alberta and southwestern Saskatchewan. Technical Bulletin No. 46. Ottawa, Canada: Canadian Dominion, Department of Agriculture. 53 p. [635]
10. Coupland, Robert T. 1950. Ecology of mixed prairie in Canada. Ecological Monographs. 20(4): 271-315. [700]
11. Coupland, Robert T.; Brayshaw, T. Christopher. 1953. The fescue grassland in Saskatchewan. Ecology. 34(2): 386-405. [701]
12. Coupland, Robert T.; Johnson, R. E. 1965. Rooting characteristics of native grassland species of Saskatchewan. Journal of Ecology. 53: 475-507. [702]
13. Cronquist, Arthur; Holmgren, Arthur H.; Holmgren, Noel H.; Reveal, James L. 1972. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Vol. 1. New York: Hafner Publishing Company, Inc. 270 p. [717]
14. Daubenmire, Rexford. 1978. Plant geography--with special reference to North America. Physiological Ecology. New York: Academic Press. 338 p. [8949]
15. Bold, H. C.; Alexopoulos, C. J.; Delevoryas, T. 1980. Morphology of plants and fungi. New York: Harper and Row. 819 p. [21159]
16. Dittberner, Phillip L.; Olson, Michael R. 1983. The plant information network (PIN) data base: Colorado, Montana, North Dakota, Utah, and Wyoming. FWS/OBS-83/86. Washington, DC: U.S. Department of the Interior, Fish and Wildlife Service. 786 p. [806]
17. Dix, Ralph L. 1960. The effects of burning on the mulch structure and species composition of grasslands in western North Dakota. Ecology. 41(1): 49-56. [808]
18. Dolan, John J.; Taylor, John E. 1972. Residual effects of range renovation on dense clubmoss and associated vegetation. Journal of Range Management. 25(1): 32-37. [12248]
19. Dolan, John Joseph. 1966. Long-term responses of dense clubmoss (Selaginella densa Rydb.) to range renovation practices in northern Montana. Bozeman, MT: Montana State University. 83 p. Thesis. [12330]
20. Douglas, George W.; Bliss, L. C. 1977. Alpine and high subalpine plant communities of the North Cascades Range, Washington and British Columbia. Ecological Monographs. 47: 113-150. [9487]
21. Eyre, F. H., ed. 1980. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters. 148 p. [905]
22. Garrison, George A.; Bjugstad, Ardell J.; Duncan, Don A.; [and others]. 1977. Vegetation and environmental features of forest and range ecosystems. Agric. Handb. 475. Washington, DC: U.S. Department of Agriculture, Forest Service. 68 p. [998]
23. Great Plains Flora Association. 1986. Flora of the Great Plains. Lawrence, KS: University Press of Kansas. 1392 p. [1603]
24. Hansen, Paul L.; Hoffman, George R. 1988. The vegetation of the Grand River/Cedar River, Sioux, and Ashland Districts of the Custer National Forest: a habitat type classification. Gen. Tech. Rep. RM-157. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 68 p. [771]
25. Hansen, Paul L.; Hoffman, George R.; Bjugstad, Ardell J. 1984. The vegetation of Theodore Roosevelt National Park, North Dakota: a habitat type classification. Gen. Tech. Rep. RM-113. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 35 p. [1077]
26. Harrington, H. D. 1964. Manual of the plants of Colorado. 2d ed. Chicago: The Swallow Press Inc. 666 p. [6851]
27. Hitchcock, C. Leo; Cronquist, Arthur; Ownbey, Marion. 1969. Vascular plants of the Pacific Northwest. Part 1: Vascular cryptograms, gymnosperms, and monocotyledons. Seattle, WA: University of Washington Press. 914 p. [1169]
28. Johnston, Barry C. 1987. Plant associations of Region Two: Potential plant communities of Wyoming, South Dakota, Nebraska, Colorado, and Kansas. 4th ed. R2-ECOL-87-2. Lakewood, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Region. 429 p. [3519]
29. Johnson, P. L.; Billings, W. D. 1962. The alpine vegetation of the Beartooth Plateau in relation to cryopedogenic processes and patterns. Ecological Monographs. 32(2): 105-135. [12218]
30. Kearney, Thomas H.; Peebles, Robert H.; Howell, John Thomas; McClintock, Elizabeth. 1960. Arizona flora. 2d ed. Berkeley, CA: University of California Press. 1085 p. [6563]
31. Kartesz, John T.; Kartesz, Rosemarie. 1980. A synonymized checklist of the vascular flora of the United States, Canada, and Greenland. Volume II: The biota of North America. Chapel Hill, NC: The University of North Carolina Press; in confederation with Anne H. Lindsey and C. Richie Bell, North Carolina Botanical Garden. 500 p. [6954]
32. Koller, Alan L.; Scheckler, Stephen E. 1986. Variations in microsporangia and microspore dispersal in Selaginella. American Journal of Botany. 73: 1274-1288. [12264]
33. Kooiman, Marianne; Linhart, Yan B. 1986. Structure and change in herbaceous communities of four ecosystems in the Front Range, Colorado, U.S.A. Arctic and Alpine Research. 18(1): 97-110. [4076]
34. Kruckeberg, A. R. 1982. Gardening with native plants of the Pacific Northwest. Seattle: University of Washington Press. 252 p. [9980]
35. Kuchler, A. W. 1964. United States [Potential natural vegetation of the conterminous United States]. Special Publication No. 36. New York: American Geographical Society. 1:3,168,000; colored. [3455]
36. Kufeld, Roland C. 1973. Foods eaten by the Rocky Mountain elk. Journal of Range Management. 26(2): 106-113. [1385]
37. Lewis, Mont E. 1970. Alpine rangelands of the Uinta Mountains: Ashley and Wasatch National Forests. Ogden, UT: U.S. Department of Agriculture, Forest Service, Region 4. 75 p. [1451]
38. Lynch, Daniel. 1955. Ecology of the aspen groveland in Glacier County, Montana. Ecological Monographs. 25(4): 321-344. [950]
39. Lyon, L. Jack; Stickney, Peter F. 1966. Two forest fires: and some specific implications in big-game management. Proceedings, Annual Conference of Western Association of Game and Fish Commissioners. 46: 181-193. [17169]
40. Martin, Alexander C.; Zim, Herbert S.; Nelson, Arnold L. 1951. American wildlife and plants. New York: McGraw-Hill Book Company, Inc. 500 p. [4021]
41. Moss, E. H. 1959. Flora of Alberta. Toronto: University of Toronto Press. 546 p. [8948]
42. Munz, Philip A. 1973. A California flora and supplement. Berkeley, CA: University of California Press. 1905 p. [6155]
43. Peck, Morton E. 1941. A manual of the higher plants of Oregon. Portland, OR: Binfords & Mort. 800 p. [12444]
44. Peet, Robert K. 1981. Forest vegetation of the Colorado Front Range: composition and dynamics. Vegetatio. 45: 3-75; 1981. [1867]
45. Ralston, R. D.; Dix, R. L. 1966. Green herbage production of native grasslands in the Red River valley. Proceedings of the North Dakota Academy of Science. 20: 57-66. [5413]
46. Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford: Clarendon Press. 632 p. [2843]
47. Reed, John F. 1952. The vegetation of the Jackson Hole Wildlife Park, Wyoming. The American Midland Naturalist. 48(3): 700-729. [1949]
48. Riggs, Robert A.; Peek, James M. 1980. Mountain sheep habitat-use patterns related to post-fire succession. Journal of Wildlife Management. 44(4): 933-938. [4546]
49. Roemer, H. L; Ogilvie, R. T. 1983. Additions to the flora of the Queen Charlotte Islands on limestone. Canadian Journal of Botany. 61(10): 2577-2580. [12217]
50. Ross, Robert L.; Hunter, Harold E. 1976. Climax vegetation of Montana: Based on soils and climate. Bozeman, MT: U.S. Department of Agriculture, Soil Conservation Service. 64 p. [2028]
51. Rowe, J. S. 1969. Lightning fires in Saskatchewan grassland. Canadian Field-Naturalist. 83: 317-324. [6266]
52. Ryerson, D. E.; Taylor, J. E.; Baker, L. O.; [and others]. 1970. Clubmoss on Montana rangelands: Distribution, control, range relationships. Bulletin 645. Bozeman, MT: Montana State University, Montana Agricultural Experiment Station. 116 p. [10855]
53. Spence, John R.; Shaw, Richard J. 1983. Observations on alpine vegetation near Schoolroom Glacier, Teton Range, Wyoming. The Great Basin Naturalist. 43(3): 483-491. [8265]
54. Taylor, John Edgar. 1967. Range pitting and nitrogen fertilization on mixed prairie rangeland in northern Montana. Bozeman, MT: Montana State University. 71 p. Thesis. [12328]
55. Tryon, Rolla M., Jr. 1955. Selaginella rupestris and its allies. Annals of the Missouri Botanical Garden. 42(1): 1-95. [11036]
56. USDA Natural Resources Conservation Service. 2018. PLANTS Database, [Online]. U.S. Department of Agriculture, Natural Resources Conservation Service (Producer). Available: https://plants.usda.gov/. [34262]
57. Van Dyne, G. M.; Vogel, W. G. 1967. Relation of Selaginella densa to site, grazing, and climate. Ecology. 48(3): 438-444. [2419]
58. Vogel, W. G.; Van Dyne, G. M. 1966. Vegetation responses to grazing management on a foothill sheep range. Journal of Range Management. 19: 80-85. [12263]
59. Wagner, Stephen Francis. 1966. Selaginella densa Rydb. and its chemical control. Bozeman, MT: Montana State University. 61 p. Thesis. [12329]
60. Weaver, J. E.; Albertson, F. W. 1956. Grasslands of the Great Plains. Lincoln, NE: Johnsen Publishing Company. 395 p. [2463]
61. Webster, Terry R.; Steeves, Taylor A. 1964. Observations on drought resistance in Selaginella densa Rydb. American Fern Journal. 54(4): 189-196. [12265]
62. Welden, Charles. 1985. Structural pattern in alpine tundra vegetation. American Journal of Botany. 72(1): 120-134. [8267]
63. Welsh, Stanley L.; Atwood, N. Duane; Goodrich, Sherel; Higgins, Larry C., eds. 1987. A Utah flora. The Great Basin Naturalist Memoir No. 9. Provo, UT: Brigham Young University. 894 p. [2944]
64. Whitman, W. C. 1979. Analysis of grassland vegetation on selected key areas in southwestern North Dakota. REAP [Regional Environmental Assessment Program] Contract No. 7-01-2. Fargo, ND: North Dakota State University, Department of Botany. 199 p. [3321]
65. Smoliak, S. 1960. Effects of deferred-rotation and continuous grazing on yearling steer gains and shortgrass prairie vegetation of southeastern Alberta. Journal of Range Management. 13: 239-243. [28733]
66. Smoliak, S. 1965. Effects of manure, straw and inorganic fertilizers on Northern Great Plains ranges. Journal of Range Management. 18(1): 11-15. [28734]
67. Wilson, Scott D.; Shay, Jennifer M. 1990. Competition, fire and nutrients in a mixed-grass prairie. Ecology. 71(5): 1959-1967. [12305]
FEIS Home Page