| FEIS Home Page |
 |
|
Photo © Thomas B. Denholm, New Jersey Department of Agriculture, Bugwood.org |
AUTHORSHIP AND CITATION:
Gucker, Corey L. 2009. Heracleum mantegazzianum.
In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station,
Fire Sciences Laboratory (Producer). Available:
https://www.fs.usda.gov/database/feis/plants/forb/herman/all.html
[].
FEIS ABBREVIATION:
HERMAN
NRCS PLANT CODE [73]:
HEMA17
COMMON NAMES:
giant hogweed
giant cow parsnip
TAXONOMY:
The scientific name of giant hogweed is Heracleum mantegazzianum
Sommier & Levier (Apiaceae) [20].
Hybrids: In several European countries, low
frequencies of giant hogweed × eltrot (H. sphondylium)
hybrids occurred in areas occupied by both parent species. Artificial
hybrids were easily created, so researchers suspect that selective
insect foraging may limit the occurrence of natural hybrids [15,63].
In North America, researchers suspect that giant hogweed × common
cowparsnip (H. maximum) hybrids are possible, but none have been
reported [42]. Although giant hogweed × eltrot hybrids are possible
in North America, none were reported in the available literature (2009). Hybrids
are discussed more in the Botanical description,
Vegetative regeneration, and
Control sections of this review.
SYNONYMS:
None
LIFE FORM:
Forb
FEDERAL LEGAL STATUS:
Noxious weed [72]
OTHER STATUS:
Information on state-level noxious weed status of giant hogweed in the United States is available from
Plants Database.
GENERAL DISTRIBUTION:
Giant hogweed is a nonnative species currently (2009) restricted to the
western and eastern parts of North America. In western North America,
giant hogweed occurs in British Columbia, Washington, and Oregon.
In eastern North America, giant hogweed occurs from Newfoundland and Nova
Scotia west to Ontario and Wisconsin and south to Indiana, Maryland,
and New Jersey ([35,36,42], NAPIS as cited in [13]). Giant hogweed is
native to the western Caucasus Mountains of Georgia, Azerbaijan, and
southern Russia [42].
In 2003, researchers reported that the range of giant hogweed was expanding in Washington, Oregon, and the northeastern United States [11]. In 1992, giant hogweed occurred in 9 Washington counties [60]. As of 2008, giant hogweed populations in Oregon were limited to areas surrounding Portland and Salem [40]. Giant hogweed is widespread in southwestern Ontario, and localized populations occur in southern Quebec, New Brunswick, Cape Breton, and eastern Newfoundland [42]. In New York, giant hogweed occurs in western and central portions of the state [14]. As of 1996, about 40 small giant hogweed populations occurred around Ithaca (Cope, personal communication, as cited in [76]). Scattered giant hogweed populations occur throughout Massachusetts [26]. Plants Database provides a distributional map of giant hogweed. As of this writing (2009), however, the map did not include Wisconsin, Indiana, Ohio, Maryland, New Jersey, Massachusetts, Vermont, New Hampshire, Nova Scotia, or Newfoundland as states or provinces with giant hogweed populations, although several sources [13,35,36,42] indicate that it occurs in these areas.
Introduction to North America: Giant hogweed was likely introduced to North America as a garden "curiosity" because of its extremely large size and impressive flower [42,74] but could have been introduced through spice importation, since its seeds are used in Middle Eastern cuisine [35]. Giant hogweed occurred in North America as early as 1917 [13].
Giant hogweed source populations in western North America were likely planted by horticulturalists [40,48]. On southeastern Vancouver Island, giant hogweed was first collected in 1964 [42], but reports of giant hogweed in southwestern British Columbia were published in the 1930s (Federation of British Columbia Naturalists Newsletter 1978, as cited in [8]). Giant hogweed has been present in Seattle, Washington, since the early 1950s (Hitchcock, personal communication, as cited in [33]). Using details about current giant hogweed habitats in western Washington and northwestern Oregon, researchers predicted that disturbed riparian habitats in the Cascade, Sierran Steppe, and Rocky Mountain ecoregions are susceptible to giant hogweed invasion [43].
In eastern Canada and the United States, giant hogweed was likely introduced as a horticultural plant [23,42]. Giant hogweed was introduced to New York state in about 1917 [23], and reports suggest that it occurred in southern Ontario by the late 1940s or early 1950s [33,74]. As of 1975, giant hogweed populations along Ontario's Saugeen River were described as "firmly established" and "very much at home" [32]. Giant hogweed was first collected from Nova Scotia in 1980 and from Quebec in 1990 [42]. It was first reported in Michigan in 1991, and the nearest known source population was in Ontario [5]. Giant hogweed was first reported in New Brunswick in 2000 [42], in Vermont in 2002, in Maryland in 2003, and in Indiana in 2004 (NAPIS, as cited in [13]).
Local distribution changes: Several studies indicate that giant hogweed is expanding its range in western and eastern North America [11]. Giant hogweed established and spread in a relatively short period of time on Vancouver Island, British Columbia. Within 40 years of its introduction, it spread along a 7.5-mile (12 km) stretch of French Creek. In some areas, populations occur 260 to 330 feet (80-100 m) from the creek's edge [9]. Over 4 years of observations in Seattle, Washington, giant hogweed populations increased in size and number each year [75]. As of 1990, the spread of giant hogweed in New York has been described as "slow but steady" (Hahn, personal communication, as cited in [75]).
In-depth studies of giant hogweed's spread have been conducted in
several nonnative European habitats. In the United Kingdom, giant
hogweed was planted in the Kew Botanical Garden in 1817 and was
first reported as an escaped ornamental in 1828. In the Czech Republic,
giant hogweed established in a noncultivated area 15 years after its introduction
in 1862 [19,50,54]. Giant hogweed experienced a lag phase that lasted over
80 years in the Czech Republic. By 1943, though, spread was exponential. Major
waterways were important to the early spread of giant hogweed. Although 90% of
spread was from riparian source populations, spread was more
rapid in nonriparian than riparian habitats (P<0.01) [49,56].
Throughout the West Bohemia area of the Czech Republic, researchers estimated
giant hogweed spread rates from aerial photos taken between 1947 (before
giant hogweed establishment) to 2000. The average aerial spread rate
was 1,261 m²/year and ranged from 139 m² to 3,275 m²/year.
The average linear spread rate was 10.8 m/year and ranged from 3.8 m
to 26.7 m/year. Because linear and aerial spread rates were not highly
correlated, researchers suggested that long-distance dispersal events
were important to the spread of giant hogweed [34].
HABITAT TYPES AND PLANT COMMUNITIES:
In its native western Caucasus range, giant hogweed occurs primarily
in meadows, clearings, or forest edges (Mandenova 1950, as cited in [22]).
Giant hogweed is common in "alluvial softwood forests" in
foothills and valley bottoms, forest clearings and abandoned grasslands
in the montane zone, and tall herbaceous vegetation in the subalpine zone
of the Western Greater Caucasus region [68].
In its nonnative North American range, giant hogweed is most common in riparian areas, open woodlands, and abandoned pasture or agricultural lands [11,28,36,40]. On Vancouver Island, initial giant hogweed establishment occurred on alluvial stream banks but soon populations spread into adjacent woodlands (personal observations by Wall and Page, cited in [42]). Past land use, disturbances, shading, and other site characteristics are likely more important than habitat type in determining a site's susceptibility to invasion. For more on these topics, see the Successional Status and Site Characteristics sections.
 |
 |
|
Photo © USDA APHIS PPQ Archive, USDA APHIS PPQ, Bugwood.org |
Photo © Thomas B. Denholm, New Jersey Department of Agriculture, Bugwood.org |
GENERAL BOTANICAL CHARACTERISTICS:
Aboveground description: Giant hogweed is a very large, monocarpic perennial forb that can reach 20 feet (5 m) tall [14,74]. While some references suggest that plants can flower at 2 years old [13,36], field research indicates that the earliest flowering age is 3 years [3,47]. The oldest plant observed in flower on an extremely dry site in the Czech Republic was 12 years old [45]. The majority of research indicates that giant hogweed dies after flowering, although one researcher reportedly observed a postflowering sprout in southern Ontario [33]. Giant hogweed × eltrot hybrids flower for mutliple years [63]. Potential nonmonocarpic behavior is discussed in Vegetative regeneration. For more on flowering, see Seed production.
Giant hogweed produces a single, hollow stem that may be 6 inches (15 cm) wide at the base [36,42,70]. Stems are distinguished from other cowparsnip (Heracleum) species by their purple blotches [14,40] and blisters or bumps with erect hairs [40]. Giant hogweed leaves are alternate, 3-part compound, and enormous. Lower leaves measure up to 10 feet (3 m) long and 5.6 feet (1.7 m) wide. Leaf size decreases up the stem [11,13,14]. At the flowering stage, plants typically have 4 to 6 stem leaves and 3 to 4 basal leaves [70]. Flowers are arranged in compound umbels that may reach 2.5 feet (0.8 m) in diameter [13,40]. Flowers open first on the terminal umbel. Flowers in the outer umbellets open first, and opening progresses inward [63]. Terminal umbels produce perfect flowers, while most flowers on lateral or secondary umbels are male [42,47,63]. Although lower umbels produce mostly male flowers, a review reports that researchers in West Bohemia observed female flower production on late-developing umbels on short basal shoots [47]. Giant hogweed fruits are dry schizocarps measuring 6 to 18 mm long and 4 to 10 mm wide [42].
Belowground description: Typically giant hogweed produces a thick, branched taproot and tightly clustered, fibrous roots [7,45]. Roots extend 18 to 24 inches (45-60 cm) deep. Lateral roots are often finer than vertical roots. Root thickness and branching increase with plant age, resulting in a "multi-taproot system" [70]. The root crown can grow to 6 inches (15 cm) in diameter and becomes lignified when giant hogweed reaches flowering age. Root crowns are often found up to 5 inches (12 cm) below ground ([70], Perrier 2001 as cited in [13]). Root crowns have contraction rings capable of shrinking the area between the crown and the shoot and protecting the hypocotyl from extreme temperatures [41].
Research in Ireland showed that giant hogweed's root system may be modified on shallow soils. In shallow soils, plants lacked a tap root and developed a laterally spreading root system. For information on the effects of shallow soil on plant growth and seed production, see Seed production.
| Important note: The sap produced by giant hogweed is toxic and causes photodermatitis. When skin is exposed to sap and the sun, severe skin rashes, blisters, and possibly permanent scarring or staining can occur [48,74]. Giant hogweed sap in the eyes can cause temporary or possibly permanent blindness [35]. Sap is considered most toxic when the plant is in flower [7]. If skin contacts giant hogweed, exposed areas should be sheltered from sunlight and washed with cold water. A physician should be consulted [40]. Burns or blisters often appear within 24 to 48 hours of plant contact [36]. | 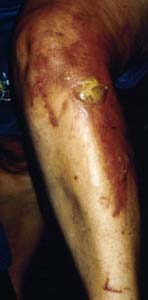 |
Photo © USDA APHIS PPQ Archive, USDA APHIS PPQ, Bugwood.org |
Raunkiaer [58] life form:
Hemicryptophyte
SEASONAL DEVELOPMENT:
Very few studies reported on the seasonal development of giant
hogweed in North America, but seasonal development is described
for several nonnative European habitats, which may relate to
similar North American climates. In Canada, first-year growth
of giant hogweed is considered "slow", but in subsequent
years growth begins earlier. In areas with mild winters, such
as British Columbia, growth may begin as early as late December.
Plants are thought to flower when they have reached a
threshold root size. These plants generally have the greatest leaf
development in the spring [42]. Flowering dates in Oregon are mid-May to July
[40] and in New England are June to July [28].
In Ireland, giant hogweed growth and development were monitored
on 7 sites. Maximum stem and leaf growth occurred from mid-May to
mid-June, peak flowering occurred in late June, and most seeds had
ripened and dispersed by late August [3]. Similar development was
reported in a review from the United Kingdom. Overwintering plants
began to grow in December. Basal leaves spread up to 20 inches (50 cm)
by March, rosettes were 3 to 7 feet (1-2 m) tall by May, flower production
peaked in early July, and plants senesced in September or October
[70]. In the West Bohemia area of the Czech Republic, giant hogweed had
flowers for an average of 36 days and a maximum of 60 days. Flowering
began in mid- to late June and was highly synchronous. Within a week,
some plants at all 10 monitored sites were flowering [46]. Roots of
flowering plants died in mid-summer and by late summer were almost
entirely decayed [45].
REGENERATION PROCESSES:
Giant hogweed reproduces by seed, and plants die after flowering.
Exceptions to this regeneration process are discussed in
Vegetative regeneration.
Pollination and breeding system: Giant hogweed flowers are self compatible and pollinated by insects [13]. In an experimental garden in Pruhonice, Czech Republic, fruit production, which ranged from 87.7% to 92.0%, was not significantly different between open-pollinated, manually outcrossed, and manually self-pollinated flowers. Nearly 4% of flowers protected from pollinators set fruit (Perglova, unpublished data, cited in [47]). During controlled studies on giant hogweed plants collected in southeastern Scotland, just 1% of flowers that were self-pollinated within the primary umbel set seed. In the primary umbel, protandry restricted the success of self fertilization. Self-pollination between flowers in the primary umbel and flowers in lower order umbels produced 68% seed set. An average of 64% of cross-pollinated flowers produced seed [63]. Field and garden studies conducted in West Bohemia revealed that male and female flowering phases overlapped for 99% of plants, suggesting that natural self fertilization is likely [46].
Although a review reports that giant hogweed's small, closely arranged, nectar-bearing flowers could be pollinated by any insect visitor [47], the most frequent insects on plants on southern Scotland's River Teriot were Coleopterans, Dipterans, Hemipterans, and Hymenopterans. Forty-eight insect species were caught on giant hogweed flowers [15]. In abandoned grasslands in southern Sealand, Denmark, giant hogweed received nearly 9 insect visits/umbel in 10 minutes of observations. Honeybees (Apis mellifera) were the most common visitors [38].
Seed production: Giant hogweed is a prolific seed producer, but reports of a single plant producing 100,000 seeds [3] are greatly exaggerated [13,47]. In western Caucasus and the Czech Republic, giant hogweed seed production ranged from 10,000 to 46,470 seeds/plant [45,46]. On the River Ayr in western Scotland, the maximum number of flowers on a single large giant hogweed plant was 81,500, and the maximum number of seeds produced by a single large plant was 52,800 [71]. It is likely that reports of 100,000 seeds produced per giant hogweed plant were calculated by doubling the number of flowers produced. Although each female flower is capable of producing 2 seeds, plants also produce a large number of male flowers. Counting all flowers as potential seeds vastly overestimates seed production [47]. Although plants do not produce 100,000 seeds/plant, giant hogweed does have an "enormous reproductive capacity" [46].
Plant age: Giant hogweed plants typically do not flower until 3 years of age. Of 302 flowering giant hogweed plants in western Caucasus and in the Czech Republic, all were at least 3 years old before producing seed. On an extremely dry site in the Czech Republic, flowering was delayed and the majority of plants flowered at 5 to 8 years of age. A single plant reached 12 years old before flowering. Flowering age differed between native and nonnative habitats and with past land use. For more information, see Successional Status.
Site conditions/plant growth: Several studies indicate that as giant hogweed height increases so does seed production. Because plant growth is often dictated by site conditions, relationships between site conditions and seed production should be expected. In Switzerland, giant hogweed plants in lowland habitats were taller and produced about twice as many flowers as those in montane or subalpine habitats (Jakob and others 1998, as cited in [41]). On field sites in Ireland, plant height and seed production were positively correlated (P<0.05). On sites where soil depths were 6 inches (15 cm) or less, giant hogweed averaged 59 inches (149 cm) tall, and seed production averaged 13,884 seeds/plant. On sites with "no soil depth restriction", plants averaged 110 inches (278 cm) tall, and seed production averaged 42,068 seeds/plant. Differences in seed production between the shallowest soil and deepest soil sites were significant (P<0.001). In this area, the researcher reported that a single plant produced 107,984 seeds [3]. Other researchers [13,47] suspect that seed production was estimated by doubling flower production, which would overestimate seed production. On the River Ayr, giant hogweed plants growing in light shade (38.9% of ambient) averaged 11 feet (3.4 m) tall and produced 208 g of seeds that individually weighed 2.25 g. In deep shade (12.6% of a mbient), plants averaged 9.5 feet (2.9 m) tall and produced 101 g of seeds that individually weighed an average of 1.18 g [71]. It is important to note that heavier seeds typically germinate faster than lighter seeds, but final germination percentages of light and heavy seeds are not significantly different (see Germination).
In West Bohemia, giant hogweed flower production was related to plant diameter and flowering date. Earlier flowering dates and increased basal diameters were correlated with increased flower production (P≤0.001). Researchers suggested that pollinators and climate were also important to flower production. Population characteristics and chemical composition of the soil did not significantly affect fecundity [46].
Seed dispersal: While the majority of wind-dispersed giant hogweed seeds fall near the parent plant, long-distance dispersal is often facilitated by water, animals, and people. In western Scotland, 60% to 90% of giant hogweed seeds fell within 13 feet (4 m) of the parent plant (unpublished data, as cited in [69]). During field studies in Ireland, Caffrey [3] found that 95% of seeds fell within 33 feet (10 m) of parent plants, but a few seeds occurred up to 160 feet (50 m) from parent plants. Caffrey suspected that seed found 160 feet (50 m) from seed-producing plants was transported by people. Field observations in Germany showed that 75% of giant hogweed seeds fell within 47 inches (120 cm) of the parent plant. The maximum seed dispersal distance was 33 feet (10 m) [41].
Wind: In a wind tunnel, giant hogweed seeds dispersed a maximum distance of 31 inches (80 cm) when released from a height of 2.3 feet (0.7 m) in wind speeds of 18 feet (5.5 m)/second [6]. Based on observations made in northwestern England, researchers suspected that wind dispersed giant hogweed seed 16 feet (5 m) from the parent plant [10]. When researchers removed leaves from giant hogweed plants in the Czech Republic, plants produced seeds that were lighter than untreated plants. Assuming lighter seeds could be dispersed further by wind than heavier seeds, cutting in giant hogweed stands may affect potential dispersal and metapopulation dynamics [52]. For more on this study, see Physical or mechanical control in the Management Considerations section of this review.
Water: Giant hogweed seeds may float in water for up to 2 days, and transport in waterways and flood water is common. Clegg and Grace [6] found that giant hogweed seeds floated 1.5 to 2 days in moving water and 3 days in stagnant water before sinking. They calculated that seeds could be transported more than 6.2 miles (10 km) in a river with a surface velocity of 0.3 feet (0.1 m)/second. Moravcova (unpublished results, cited in [30]) reported that 6-month-old giant hogweed seeds sank within 8 hours in water. Importance of water to the transport of giant hogweed seed is also discussed in Spread, Impacts, and Control.
Localized winter flooding may also disperse giant hogweed seeds. Giant hogweed was eliminated from a meadow in northeastern Zealand, Denmark, after 7 years of sheep grazing. No giant hogweed seedlings emerged from soil samples collected from grazed sites, but a few seedlings emerged on grazed sites in the field. Researchers thought seed was dispersed from adjacent ungrazed sites by sporadic flooding during the previous wet winter [1]. A review reports that winter floods can detach entire giant hogweed skeletons, transporting entire umbels [70].
Animals: Animals may disperse giant hogweed seed. Although giant hogweed seeds were not taken by birds in Edinburgh, Scotland [6], researchers suggested that giant hogweed populations in Michigan may have resulted from bird dispersal, because dispersal from the nearest source population in Ontario by water was unlikely [5]. Cattle may have dispersed giant hogweed seed, based on observations of invaded sites in northwestern England. Whether seed was transported in hide, hooves, or dung was not discussed [10].
Humans: Through the collection of flowers, movement of soil, intentional plantings, and importation of seed, humans have been important in giant hogweed seed dispersal. In British Columbia, a landowner observed a couple load their truck with dried giant hogweed stalks and umbels and drive away [9]. In north Vancouver, British Columbia, soil movement was important to the introduction of giant hogweed seed (Page, personal observation, as cited in [42]). In New York, giant hogweed has been dispersed by intentional plantings and the collection of dried fruits [36]. During a survey of giant hogweed-invaded sites in northwestern England, researchers suspected seed was transported by equipment or vehicles [10]. A review reports that giant hogweed is a preferred honeybee food and that beekeepers in Europe aided in the dispersal of giant hogweed [13].
Giant hogweed seed is often intercepted by Animal and Plant Health Inspection Service personnel. Seeds are often brought to North America by Middle Eastern tourists, who use the seeds as a spice (Westbrooks and Preacher 1986, as cited in [76]). At Sea-Tac airport, giant hogweed is one of the most frequently intercepted noxious weed seeds. Seed is typically found in the personal baggage of passengers, especially those from Iran [62]. Importance of human activities in the dispersal of giant hogweed is also discussed in the Spread, Impacts, and Control section.
Seed banking: The majority of giant hogweed seeds germinate within a year of their production; however, a small portion may persist in the soil at least 3 years and perhaps as long as 7 years. A review reports that at least one giant hogweed seed was viable after 15 years of indoor storage, but that in the field seeds did not germinate after 7 years in the soil [13]. After 7 years of sheep grazing in a meadow in Zealand, Denmark, giant hogweed was eliminated from the grazed area and no seedlings emerged from soil samples collected from the grazed sites. Prior to their elimination, plants on grazed sites did not set seed, suggesting that the giant hogweed seed bank may be depleted in less than 7 years [1].
Giant hogweed seeds collected in western Czech Republic and stored in dry conditions for up to 3 years germinated at a significantly lower percentage than fresh seed (P<0.001). For all temperatures tested, 30% to 93% of fresh seed germinated. After a year of storage, 2% to 41% of seeds germinated, and after 3 years of storage, 0% to 9% germinated. In a field experiment, an average of 8.8% of buried giant hogweed seed germinated after a year in the soil, 2.7% germinated after 2 years in the soil, and 1.2% germinated after 3 years in the soil. Researchers noted that, given giant hogweed's prolific seed production, even a low percentage of seed survival would allow for some establishment [31]. After 1 year of burial, most nongerminating seeds had fully developed embryos, although some had only partially developed embryos [30].
Generally, giant hogweed seeds are abundant in the soil near flowering plants. The majority of seeds occur in the top 2 inches (5 cm) of soil. When soil samples were collected from up to 6-inch (15 cm) depths in the Czech Republic, 95% of giant hogweed seeds were recovered from the top 2 inches (5 cm) [22]. A review reports that in a large giant hogweed stand in Germany there were as many as 2,664 giant hogweed seeds per m² [70]. In the Czech Republic, the seed bank was nearly depleted with spring germination. Viable seed density averaged 3,759 seeds/m² in the fall, 2,044 seeds/m² in the spring, and 192 seeds/m² in the summer. Researchers estimated there were 386 million nondormant seeds at the time of spring germination in the largest giant hogweed population, which occupied 1,066,900 feet² (99,121 m²) [22].
Germination: Giant hogweed seeds require cold, moist stratification [30], and early spring germination is most common in giant hogweed's nonnative range [3,36,42,44]. High germination percentages are reported from laboratory studies [29,42]. A review reports that germination can occur in the dark, and that giant hogweed germinates well on recent deposits along river banks, in organic matter in woodlands, and in open areas with "adequate" light and moisture [70].
In laboratory tests, giant hogweed germination percentages were as high as 91% [29]. Fifty to eighty percent of seeds collected from overwintering stems in Ontario and Quebec germinated at room temperature in the laboratory (Darbyshire, unpublished data, cited in [42]). For 1,680 seeds collected in the western Czech Republic, germination averaged 91.1% in the laboratory. Germination percentages were not related to seed size or position on the plant (terminal, lateral, or satellite umbels). The largest seeds were produced in terminal umbels and germinated faster than small seeds, but final germination percentages were not different between large and small seeds [29].
In British Columbia, giant hogweed seeds germinate in late winter or early spring (Wall, personal observation, as cited in [42]). In New York, seed germination begins in early spring but may continue throughout the growing season [36]. On 2 sites in Ireland, giant hogweed first emerged in February. Emergence peaked in late April and decreased thereafter [3]. In West Bohemia, the majority of germination occurred after snowmelt, generally in April. No new giant hogweed seedlings emerged after May, and there was no germination in the fall (Pergl and others, unpublished information, as cited in [44]). Fall germination has been observed by researchers in Poland, Ireland, and Scotland [44].
Seeds from cut umbels may still germinate [52]. This is important when considering potential control methods and site clean-up. For more on this topic, see the Spread, Impacts, and Control section.
Stratification: Giant hogweed seeds exhibit morphophysiological dormancy. Embryos of ripe seeds are not fully developed, and dormancy is broken by cold, wet conditions [30]. Almost no seedlings emerged from soils collected in October in northeastern Zealand, Denmark. Emergence was much greater from soils collected in March, suggesting cold temperature exposure increased germination [1]. In the Czech Republic, 99.7% of seeds shed in the fall were dormant, but by spring, only 12% of seeds were dormant [22]. Fresh giant hogweed seeds collected from the Sheffield region of England did not germinate in the laboratory. After 2 months of cold, moist stratification, 40% of seeds germinated. Germination percentages between stratified and fresh seed were significantly different (P<0.001) [16]. A review reports that giant hogweed seeds failed to germinate when dried before exposure to cold temperatures [70].
Temperature: After giant hogweed seeds collected in the Czech Republic were stratified, germination percentages were greatest (93%) at alternating temperatures of 41 and 68 °F (5 and 20°C). At a constant temperature of 43 °F (6 °C), germination was 88%. Germination was poorest at a constant temperature of 72 °F (22 °C). Seeds at low constant temperatures germinated slowly over a long period of time. Stratification was typically successful after 2 months or more at temperatures below 50 ° F (10 °C), and dormancy returned with increasing temperature. Researchers predicted that giant hogweed may produce a fraction of seeds that require multiple seasons to break dormancy [30,31].
Seedling establishment and plant growth: Research and observations suggest that giant hogweed establishment is more likely on open or disturbed sites than on sites with established vegetation, especially grasses. Generally, seedling survival is low and may be even lower on shaded or flooded sites. Pergl and others [44] report in a review that the probability of giant hogweed establishment is very low in undisturbed grasslands and other vegetation types, but that in already invaded sites, the dying parent provides an open site for establishment. In Ireland, giant hogweed establishment is typically greatest on previously disturbed sites [4]. In mid-summer in a dense grassland in the Czech Republic, there were 56 giant hogweed seedlings on a 1-m² plot next to an adult plant that flowered and died in the previous season (Pergl and Perglova, unpublished work, as cited in [44]). In the Czech Republic, the density of giant hogweed seedlings was not related to adult plant density or seed bank density (Pergl, unpublished work, as cited in [44]).
Survival: The percentage of giant hogweed seedlings surviving an entire year is often low. Reports on seedling survival in North America were lacking as of this writing (2009). During field monitoring of 2 Ireland sites, 1.2% to 13.7% of giant hogweed seedlings survived until August of the emergence year [3]. In Germany, survival of giant hogweed seedlings was less than 1% (Huls 2005, as cited in [44]), and from March to June in Scotland, the survival of giant hogweed seedlings and immature plants was about 2.5% (Caffrey 2001, cited in [44]). In the Czech Republic, less than 1% of giant hogweed seedlings survived 1 year. The probability of giant hogweed seedling survival to the next year increased with increasing leaf number and length of the longest leaf (Pergl, unpublished work, as cited in [44]).
Flooded or shaded conditions may decrease the survival of giant hogweed seedlings. Seedlings along a river in Germany did not survive summer flooding [65]. In Scotland, 400 giant hogweed seedlings/m² occurred soon after seed bank germination, but density dropped to 33 seedlings/m² beneath the shading of giant hogweed adults present by mid-summer [70].
Growth rate: Reports about the growth rate of giant hogweed seedlings are conflicting [4,11,70]. Without more information, it is difficult to know if these reports conflict because of regional differences or because of different reference points. In a laboratory study, giant hogweed seedlings grew 0.155 to 0.186 g/g/day. Seedlings grown from seed collected in the terminal umbel grew most rapidly. The relative growth rate of giant hogweed seedlings ranked 34th highest among 117 herbaceous species occurring in the United Kingdom. Giant hogweed seedlings grew at rates similar to those of grasses [44].
Vegetative regeneration: Most research indicates that giant hogweed dies after flowering and that vegetative regeneration is restricted to damaged plants or hybrids. During extensive studies of giant hogweed in its native Caucasus habitats and in its nonnative habitats in the Czech Republic, all plants were "strictly monocarpic" [45]. In southern Ontario, however, field observations led Morton [33] to suspect that some giant hogweed plants sprouted after flowering. European researchers have suggested that Morton mistook a seedling near a dead stem for a vegetative sprout but acknowledged that perennial species occur within the Heracleum genus and suggested that a perennial giant hogweed growth form may be possible [45,47].
It is generally accepted that giant hogweed hybrids may be perennial and that sprouting occurs when flowering plants are damaged. Giant hogweed × eltrot hybrids, produced when eltrot was the maternal parent, reproduced successfully and did not die after flowering. Hybrids are likely short-lived, however. Naturally occurring hybrids found in southern Scotland in 1977 were not found in 1982 [63]. Damaged giant hogweed plants may sprout following damage and survive until their flowering year [70]. In late June in Germany, giant hogweed cut at the base flowered within 3 weeks of cutting [41]. For more on the survival of damaged plants, see Spread, Impacts, and Control.
SITE CHARACTERISTICS:
Descriptions of giant hogweed habitats in North America are less
detailed than those of its nonnative European habitats. Characteristics
of invaded European habitats may provide information about the
susceptibility of habitats in North America, where giant hogweed
is still expanding its range.
Giant hogweed is often described along roadways and waterways in North America, which have been important in the early spread of giant hogweed in Europe (see Spread, Impacts, and Control). Along French Creek on Vancouver Island, giant hogweed initially established on alluvial stream bank soils but more recently, giant hogweed has spread into adjacent woodlands (Wall, personal observation, as cited in [42]). This same pattern was noted in northern Vancouver (Page, personal observation, as cited in [42]). In Washington and Oregon, giant hogweed occurs in riparian areas, disturbed sites, and along roads [11]. In the eastern Midwest, giant hogweed habitats include roadsides, stream banks, railway embankments, and abandoned fields [7]. Giant hogweed occurs in the sandy gravel along roadsides and ditches in eastern Ontario and western Quebec (Darbyshire, personal observation, as cited in [42]). Open woodlands, canals, streams, roadsides, and vacant lots support giant hogweed populations in New England and New York [28,36].
In Europe, giant hogweed is common in unplowed pastures, meadows, riparian areas, and forest clearings in high-humidity areas with nitrogen-rich, basic soils [61], although many soil types are tolerated. During a survey of giant hogweed-invaded habitats in northwestern England, none of the habitat charactersitics measured (altitude, slope, distance from water, bank top width, and water quality) consistently predicted giant hogweed's occurrence. Researchers proposed that either habitat characteristics were too broad or giant hogweed's tolerances too general for good prediction and concluded that "almost all United Kingdom banksides (we)re susceptible to invasion" [10]. Using logistic regression modeling, researchers found that human population density (P<0.0001), low winter temperatures (P=0.002), and moraine clay (P=0.001) significantly predicted the distribution of giant hogweed on a national scale in Denmark. At the regional scale within Denmark, predictive variables were human population density (P<0.0001), alluvial sand cover (P=0.001), and spring precipitation (P=0.01) [37]. In West Bohemia, researchers comparing invaded and uninvaded sites found that giant hogweed did not occur in vegetation types with acidic soils, including peat bogs, birch (Betula spp.) woodlands, and spruce (Picea spp.) forests. Habitat susceptibility increased with increased light availability (P=0.0117), increased soil reaction indicators (P=0.0004), increased dispersal potential (P<0.001), and increased disturbance (P<0.001) [57].
Climate: The climates in giant hogweed's native and nonnative ranges are similar. In the Western Greater Caucasus region, giant hogweed occupies habitats from 160 to 7,200 feet (50-2,200 m) [68]. In this area, January temperatures can range from -33 to 61 °F (-36 to 16 °C) and July temperatures from 27 to 95 °F (-3 to 35 °C). Based on 30 years of data, annual precipitation averaged 34 inches (863 mm) [45]. Several reviews have summarized the prevailing climates in giant hogweed's nonnative range. Plants generally grow best in northern temperate climates [70]. Cold winters are common in giant hogweed-invaded habitats and are likely necessary for successful germination and establishment (Tiley and others 1996, as cited in [13]). Giant hogweed seedlings are frost resistant (Godefroid 1998, as cited in [42]). While moist sites are preferred, moderate summer droughts are also tolerated [70]. In Canada, giant hogweed is considered "most invasive" in cool, moist climates, and populations occur as far north as Kapuskasing, Ontario. Giant hogweed is also found on seashores and tolerates salt spray [42].
During population and growth studies conducted in Germany, giant hogweed suffered high mortality during a hot dry summer, but reproduction and recruitment from drought survivors was high [18]. In the Czech Republic, giant hogweed's distribution was significantly related to January isotherms (P=0.003). Giant hogweed was less common in areas with warm winters. Average June isotherms, annual precipitation, and elevation did not significantly affect giant hogweed's distribution [51]. Distribution was also affected by human population density; for more about this see Spread, Impacts, and Control. When germination, establishment, and growth of giant hogweed were evaluated along elevation gradients in the United Kingdom and the Czech Republic, researchers found that limitations to dispersal explained giant hogweed's distribution better than climate. At all sites tested from 30 to 2,000 feet (10-600 m), more than 80% of giant hogweed seed germinated. Second-year survival of giant hogweed was lowest at the highest-elevation, coldest-climate site but was still almost 30% [77].
Soils: In most reviews, giant hogweed is described on deep, moist soils with nearly neutral pH and moderate to high nutrient content. Soil textures in giant hogweed habitats may be gravels, sands, loams, or clays. Moist soils are preferred, and waterlogged and winter-flooded soils are tolerated but extended periods of inundation are not [13,28,42,70]. Giant hogweed seeds rot quickly on sites inundated for long periods [70]. The pH reported from giant hogweed habitats ranged from 4 to 8.5 [68].
In southern Ontario, giant hogweed is often found on deep, rich, moist soils [33]. In Germany, giant hogweed was most common on loamy soils. Giant hogweed-invaded sites had an average pH of 5.6, experienced rapid nutrient cycling, were well aerated, and received a "good supply" of water [65]. In Edinburgh, Scotland, giant hogweed populations occurred on soils with pH levels ranging from 5 to 9 and organic matter contents ranging from 3.1% to 8.7% [6].
SUCCESSIONAL STATUS:
Most studies and observations indicate that giant hogweed establishes best in
recently disturbed sites dominated by early-seral vegetation. After
establishment, populations may spread into more successionally advanced vegetation
types. In giant hogweed-invaded sites, full sunlight is common, although giant
hogweed also occurs in shaded habitats. Although common on disturbed
sites, giant hogweed abundance typically decreases with tilling and
repeated mowing. Giant hogweed may grow best on sites that were disturbed
prior to its establishment but remain undisturbed after its establishment.
In Germany, the highest density of giant hogweed occurred in areas dominated by early-seral vegetation. Generally, the cover of giant hogweed decreased with increasing successional age of the site. Of the plots invaded by giant hogweed, 71% occurred in abandoned fields, pastures, grasslands, or "other derelict land". Giant hogweed densities were low in managed areas that were grazed or mowed regularly. Densities were much greater on sites where grazing or mowing had been discontinued or occurred irregularly. "Recent" mechanical disturbances occurred on 27% of invaded plots, and periodic flooding occurred on 34% of invaded plots. The number of flowering giant hogweed plants on 25-m² plots was 5 in woodlands, 26 in managed grasslands, and 37 in ruderal grasslands. Researchers noted that fruit production was "strongly reduced" on managed grasslands [65,68].
Giant hogweed population density was higher on grazed pastures than in undisturbed habitats in the Czech Republic, but plants took longer to flower in pastures than on undisturbed sites (P<0.001). In pastures, vegetative plants likely made up a large portion of the population. In giant hogweed's native Caucasus habitats, population density and flowering age were not different between pastures and undisturbed areas. Pastures in the Czech Republic were grazed 3 times/year by sheep, and in Caucasus, grazing was less extensive. The proportion of flowering plants in the youngest age class in unmanaged Czech and Caucasus sites was 89% and 31%, respectively. Differences in the flowering age between unmanaged Czech and Caucasus sites were likely related to elevation differences. Higher elevation sites in the Caucasus region likely experienced a shorter growing period [45].
| Population density and flowering age of giant hogweed in native and nonnative pasture and unmanaged habitats [45] | |||
| Habitat | Population density (individuals/m²) | Average flowering age (years) | Maximum flowering age (years) |
| Native, unmanaged | 4.38 | 4 | 7 |
| Native, pasture | 4.99 | 5 | 7 |
| Nonnative, unmanaged | 4.58 | 3 | 4 |
| Nonnative, pasture | 11.17 | 5 | 7 |
Shading: In its nonnative North American range, giant hogweed is described in full-sun, part-shade, and full-shade conditions [26,28,33]. While a wide range of shade tolerance is also reported in giant hogweed's nonnative European habitats [4,65], most studies indicate that giant hogweed is more common in full sun or partial shade. In central European beech (Fagus sylvatica) forests in Switzerland and Denmark, giant hogweed "occasionally invades" gaps created by tree fall, but with canopy closure, giant hogweed abundance decreases until it is eventually eliminated (Ravn, unpublished observations, cited in [59]). In Germany, 46% of giant hogweed-invaded sites were classified as full light, 32% as light, 15% as semi-shade, and 7% as shade. Researchers described plant growth as "fair" in partial shade. Giant hogweed did not occur beneath deeply shaded closed-canopy woodlands but did occur beneath sparse canopies, in canopy gaps, and at woodland edges. Shrubs rarely occurred in giant hogweed-invaded sites. Maximum shrub cover in giant hogweed stands was 10% [65].
Postfire regeneration strategy [64]:
Crown residual colonizer (on site, initial community)
Secondary colonizer (on- or off-site seed sources)
Fire adaptations and plant response to fire: Giant hogweed's protected root crown makes postfire sprouting likely. Root crowns often occur up to 5 inches (12 cm) below ground where they would be protected from extreme temperatures ([41,70], Perrier 2001 as cited in [13]). Seedling establishment on burned sites is also likely, given that seed germination and seedling growth are best on open and/or previously disturbed sites [4,70].
FUELS AND FIRE REGIMES:
Fire regimes in giant hogweed's native range were not described in the
reviewed literature. Pysek and Morvcova [53] report that fire is not common
in giant hogweed's native and nonnative European habitats. Giant hogweed's
preference for moist sites suggests that fires may be infrequent and burn with
low severity in its nonnative North American habitats. A complete absence of
fire in invaded North American sites, however, is unlikely. See the
Fire Regime Table for further
information on fire regimes in vegetation communities where giant hogweed may occur. Find further fire regime information for the plant communities in which this
species may occur by entering the species name in the
FEIS home page under "Find Fire Regimes".
FIRE MANAGEMENT CONSIDERATIONS:
Potential for postfire establishment and spread:
Although information about fire in giant hogweed habitats is lacking, managers
could expect giant hogweed to sprout following fire, since its recovery following stem
injury is well established [41,70]. Open, burned areas could also provide suitable
habitat for seedling establishment if a
seed source exists. Giant hogweed is dispersed by a variety of vectors
(see Seed dispersal). Burned sites in the vicinity of
established giant hogweed populations should be monitored for giant hogweed seedlings
for several years, and appropriate control measures taken.
Palatability and/or nutritional value: Information on the digestibility and nutritional quality of giant hogweed is provided in a review by Buttenschon and Nielsen [2]. Giant hogweed plants in Germany had low nitrogen and high phosphorus levels; for more details, see the paper by Otte and Franke [41].
OTHER USES:
A review reports that giant hogweed is a valuable honeybee food
source and is useful to European beekeepers [13]. Giant hogweed
seeds are also known as golpar, a spice used in Iranian cooking [35].
Both of these uses were likely important to the dispersal of giant
hogweed. For more information, see the section on seed dispersal by
humans.
SPREAD, IMPACTS, AND CONTROL:
Spread:
In Canada, giant hogweed populations generally establish along
roadsides, forest edges, and streams, but populations have spread
into adjacent open woodlands, grasslands, and meadows. Researchers
suggest that managers monitor these corridors during control
operations [42]. Similar corridors were important to the early
spread of giant hogweed in Europe. Given that giant hogweed
established over a decade earlier in Europe than in North America,
recognizing similarities between spread patterns and vulnerable habitats
in Europe and North America may improve management and control of this species.
In Edinburgh, Britain, researchers found that giant hogweed spread along rivers, railways, and roadsides. On riverbanks, giant hogweed was mixed with the riparian community. On drier and disturbed sites, giant hogweed often dominated. Researchers suspected that seeds were often dispersed downstream from gardens and established once they settled on the streambank. Dispersal away from the river was likely the result of human activity along transportation routes. Mud trapped in the boots of railway workers and the collection and transport of umbels were considered important to dispersal [6]. In Germany, giant hogweed was most frequent on riverbanks, ditches, roadsides, or paths [67]. Based on regression analyses, the occurrence of giant hogweed decreased with increasing distance from rivers (P=0.0004), increasing distance from agricultural roads (P<0.0001), and increased presence of woody vegetation (P=0.0019) [69]. Giant hogweed was most common in early-seral vegetation. This topic is discussed in Successional Status.
Linear landscape features such as roads, paths, and streams were important to the initial spread of giant hogweed in West Bohemia [34]. With time, giant hogweed became less restricted to the cool climates at high elevations. In 1970, 28.5% of giant hogweed populations were restricted to elevations above 2,000 feet (600 m). In 1990, just 14.7% of populations occurred at elevations above 2,000 feet (600 m) (Pysek 1994, as cited in [54]). When giant hogweed habitats were surveyed, 83.4% of invaded areas were treeless pastures, meadows, or fields. Forests presented a barrier to invasion and "only very rarely" did giant hogweed occur in forest interiors [34]. In the Czech Republic, giant hogweed's distribution was significantly affected by the density of the human population (P=0.01). Giant hogweed was less common in areas with fewer than 60 people/km² [51].
Impacts: In the United States, giant hogweed is a noxious weed. Contact with plant sap can burn, blister, and scar exposed skin. Large plants can restrict the use of and access to rivers, trails, and paths [40,66], and studies indicate that giant hogweed can negatively impact soil dynamics, fisheries, and other species in its nonnative habitats. In one study, however, the presence of giant hogweed had a positive influence on the pollination of an associated species. For additional information on the health effects of toxic giant hogweed sap, see the Important note in the Botanical and Ecological Characteristics section.
Soil dynamics: In Belgium and the Czech Republic, researchers found that soil nutrient content was different on giant hogweed-invaded and uninvaded sites. In central Belgium, carbon mineralization and concentrations of carbon and nitrogen were lower on invaded than uninvaded sites. Researchers suggested that giant hogweed litter may decompose more slowly than that of native species and could decrease organic matter turnover [21]. Conversely, during studies conducted in West Bohemia, researchers indicated that the high production and easy decomposition of giant hogweed biomass increased nutrient concentrations on invaded sites [57]. Direct measurements of giant hogweed decomposition rates would be a good first step to address the apparent contradiction of these studies.
Stream bank dynamics/fisheries: In the western United States, a review notes that dense giant hogweed canopies could shade and eventually replace native riparian species [11,40]. Decreased understory vegetation could increase the potential for stream bank erosion [11]. Studies in Ireland and Scotland have shown that giant hogweed skeletons leave stream banks exposed to erosion during rain or flooding events. When giant hogweed replaces the persistent stream bank graminoids and other perennials, silt deposits can increase to a degree that makes gravel substrates unsuitable for salmon spawning [3,66,71].
Other species: In nonnative European habitats, studies have reported both reduced and increased species richness on sites with giant hogweed. Studies on giant hogweed's effects on native North American habitats are lacking. A review from southern Scotland reported greater species diversity in grasslands with giant hogweed than in uninvaded grasslands. In woodland habitats, species diversity and cover were lower on invaded than uninvaded sites. Researchers suggested shading by giant hogweed may have provided improved establishment conditions for some grassland species [70]. In Germany, researchers found a negative relationship between giant hogweed cover and the number of vascular species (P<0.001, R²=0.07); however, the relationship varied by community type. Species richness was reduced most, by 4.8 species, when giant hogweed occurred in tall herbaceous communities. Researchers noted that these effects were local and that regional native species populations were not endangered [66]. In West Bohemia, giant hogweed-invaded vegetation had 40.5% fewer species than uninvaded vegetation [57].
Pollination/pollinators: In southern Sealand, Denmark, researchers conducted a field study to determine whether or not giant hogweed negatively affected the pollination of a co-occurring species. They found that seep monkeyflower (Mimulus guttatus) received more insect visits when plants were located next to giant hogweed than when plants were located at any distance 30 to 660 feet (10-200 m) away from giant hogweed (P≤0.01). More insect visits, though, did not increase seed production by seep monkeyflowers, which are capable of self pollination. Researchers suggested that giant hogweed may be important to pollinators in species-poor grasslands [38]. Pollinators attracted to giant hogweed are noted in the Pollination and breeding system section.
Control: Integrated control methods and vigilant monitoring may prove most useful in the management of giant hogweed populations. Public education, coordinated efforts, and prioritized treatments are also important to successful control. Studies on the control of giant hogweed in its nonnative European and North American habitats are presented below.
Reviews provide important general information for giant hogweed management. Public education is important to giant hogweed management, not only to minimize health hazards but also to limit transplanting and seed dispersal [42]. Mapping giant hogweed populations before implementing a control program can improve the coordination of management efforts and posttreatment monitoring. Researchers suggest that searches for giant hogweed seedlings should be conducted at all treatment sites for at least 5 years after all plants have been removed [39]. The Forestry Commission in western Europe suggests that recently established giant hogweed populations should receive management priority, and when giant hogweed occurs along a waterway, populations at the uppermost stream should be controlled first [78].
Because giant hogweed reproduces only by seed, produces a short-lived seed bank, and dies after flowering, preventing seed production and depleting the seed bank seem like viable control strategies [42,63]. However, giant hogweed roots remain alive for "many" years if plants are kept from flowering [60]. Some suggest that preventing seed spread and seed production can be more labor intensive and expensive than "aggressive" control efforts [39]. In areas where giant hogweed ×eltrot hybrids are possible, eliminating seed production is an even less viable control strategy, as hybrids often do not die after flowering [63]. The following references provide information about a variety of potentially useful giant hogweed control techniques and methods: [12,13,27].
Fire: For information on the potential use of fire in giant hogweed control efforts, see Fire Management Considerations.
Prevention: In North America, early detection of new giant hogweed populations and prevention of seed dispersal are management priorities. Hotlines exist in Oregon, New York, and New Jersey for reporting suspected giant hogweed sightings [35,36,40]. Giant hogweed seed is often intercepted by Animal and Plant Health Inspection Service personnel (Westbrooks and Preacher 1986, as cited in [76]). At Sea-Tac airport, giant hogweed is one of the most frequently intercepted noxious weed seeds. Seed is typically found in the personal baggage of passengers, often those from Iran, who use the seeds in cooking [62]. As of 2006, Canadians did not classify giant hogweed as a federal noxious weed or prevent its entry into Canada. In parts of British Columbian and Ontario, though, giant hogweed is classified as noxious [42]. As of 2005, giant hogweed was still available for sale in Canadian nurseries (Darbyshire, personal communication, as cited in [42]).
Because the probability of giant hogweed establishment is low in undisturbed grasslands and other undisturbed vegetation types, land managers in Europe attempt to restrict giant hogweed establishment and spread by minimizing disturbances in established vegetation [44].
Physical or mechanical control: Repeated mowing or cutting can reduce or eliminate giant hogweed seed production ([3], Nielsen and others 2005 as cited in [39]). Cultivation could be an effective control method on some sites [78], and hand pulling may be effective for small patches of seedlings [42].
Cutting: Belowground root cutting is the only mechanical control method that reliably killed giant hogweed plants in Scotland [71]. Aboveground stem damage rarely caused mortality but often reduced plant size and/or seed production [3]. In western Europe, plants cut 4 times a year for 2 years were not killed (Rubow 1990, as cited in [39]), but some indicate that frequent cutting may encourage the growth of grazing-tolerant vegetation and provide some competition for resources in giant hogweed stands [39]. It is important to note that stems cut during the flowering stage may still produce viable seed [52]. Some do not recommend mowing, cutting, or weed whacking giant hogweed stands, since exposure to the sap is likely, and regrowth from the taproot is rapid [36].
In western Scotland, giant hogweed plants cut at 2 inches (5 cm) below ground failed to flower, and plants cut at 6-inch (15 cm) depths were killed. Plants cut at ground level produced umbels that averaged 9.8 g (dry weight); uncut plants produced umbels that averaged 227.5 g. Plants cut farther above ground produced significantly fewer flowers and seeds than uncut plants (P<0.001) but more than plants cut below ground [71]. While root cutting may provide some control, the process is labor intensive and if done in a dense stand, will require checks for missed plants [39]. Giant hogweed plants that were cut at ground level in Ireland were not killed but were shorter and produced fewer seeds than uncut plants. Late-May cutting reduced plant heights and seed production more than early March cutting. Uncut giant hogweed plants averaged 96 inches (244 cm) tall and produced an average of 34,113 seeds/plant. Giant hogweed plants cut in late May averaged 50 inches (127 cm) tall and produced 4,281 seeds/plant [3]. Even plants cut in July flowered and produced seed [4].
Timing of cutting treatments can affect potential seed production and future control efforts. Some recommend that managers cut giant hogweed plants in the mid-flowering stage, which reduces seed production but also allows large plants to potentially shade out seedlings. Reduced seedling number should decrease control efforts necessary in the following year (Nielsen and others 2005, as cited in [39]). In the Czech Republic, the majority of plants survived leaf, flower, and leaf and flower removal treatments that occurred in early June (bud stage), mid-June (start of flowering), and early July (start of fruit development). Seeds produced by untreated plants were heavier than those of most treated plants, but lighter seeds germinated as well as or better than the heavier seeds of uncut plants. When umbels were removed in early June, 90% of plants regrew and produced an average of 3,310 fruits/plant. When umbels were removed in mid-June and in early July, 95% and 80% produced an average of 461 and 446 fruits/plant, respectively. Umbels cut from 85% of the plants produced viable seed while detached from the plant. Although germination of seeds from cut umbels was significantly lower than that from intact umbels (P=0.039), researchers calculated that an average of 442 viable seeds/cut umbel were left on treated sites. A maximum of 20%, 30%, and 50% of seeds from umbels cut in early June, mid-June, and early July germinated, respectively. Findings from this study suggest that seeds produced in cut areas may disperse farther, affecting metapopulation dynamics. Disposal of flowering stems on treated sites is important. Burning of cut stems may be the best method of disposal [52,55]. For more on this, see Fire Management Considerations.
Biological control: Control of giant hogweed by insects or pathogens may be unlikely, but several studies suggest that livestock grazing may be useful to manage giant hogweed. Insects and pathogens potentially useful in the biological control of giant hogweed are being investigated [42], but the existence of closely related native cow parsnip species makes their release unlikely [61].
Farmers near Parksville, British Columbia, noted reduced giant hogweed abundance in pastures used by cattle, goats, and/or hogs [42]. Sheep essentially eliminated giant hogweed from a meadow in northeastern Zealand, Denmark. During the grazing study, there were 5 sheep/ha from 1987 to 1989 and 10 sheep/ha from 1990 to 1993. Cover of giant hogweed was as high as 19% in May of 1989 but was 0% in May of 1993. Because a few giant hogweed seedlings emerged on grazed sites with a depleted seed bank, researchers suggested that if a nearby seed source exists, monitoring and grazing treatments should continue after the removal of giant hogweed [1]. After 4 to 5 months of livestock grazing along a stream in Denmark, the density of giant hogweed seedlings (P=0.1) and large flowering plants (P<0.05) was lower, but the density of small vegetative plants was higher on grazed than ungrazed sites (P<0.001) (Sorensen 2002, as cited in [2]).
Guidlines for the use of livestock to control giant hogweed are reported in a review by Buttenschon and Nielsen [2]. Giant hogweed may be controlled best when livestock feed on early growth and are used to deplete nutrients stored in the taproot through either continuous or recurrent grazing. In dense giant hogweed stands, heavy grazing 3 to 4 times/year may be most effective. In less-dense stands, summer grazing is recommended. Animals experienced with giant hogweed grazing can be used to teach inexperienced individuals to feed on giant hogweed and to avoid overeating, which can cause poisoning and/or digestive problems. Dark-skinned and/or densely haired livestock breeds are least susceptible to dermatitis [2].
Chemical control: Several references provide information on the use of chemicals to control giant hogweed: [4,39,78]. The Forestry Commission of western Europe suggests that herbicides are most effective when giant hogweed is growing but before plants become large, which is in April or May [78]. Herbicide treatments applied when plants are large will not likely penetrate the canopy and reach smaller plants [39].
Integrated management: In New York, small giant hogweed populations were successfully managed with repeated digging and herbicide treatments [23]. Researchers suggest that the reestablishment of grasses is important to successful control on treated sites [39].
The following table provides fire regime information in habitats where giant hogweed may occur. Find further fire regime information for the plant communities in which this species may occur by entering the species name in the FEIS home page under "Find Fire Regimes".
| Fire regime information on vegetation communities in which giant hogweed may occur. This information is taken from the LANDFIRE Rapid Assessment Vegetation Models [25], which were developed by local experts using available literature, local data, and/or expert opinion. This table summarizes fire regime characteristics for each plant community listed. The PDF file linked from each plant community name describes the model and synthesizes the knowledge available on vegetation composition, structure, and dynamics in that community. Cells are blank where information is not available in the Rapid Assessment Vegetation Model. | ||||||||
|
||||||||
| Pacific Northwest | ||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | ||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
|||||
| Northwest Grassland | ||||||||
| Bluebunch wheatgrass | Replacement | 47% | 18 | 5 | 20 | |||
| Mixed | 53% | 16 | 5 | 20 | ||||
| Idaho fescue grasslands | Replacement | 76% | 40 | |||||
| Mixed | 24% | 125 | ||||||
| Alpine and subalpine meadows and grasslands | Replacement | 68% | 350 | 200 | 500 | |||
| Mixed | 32% | 750 | 500 | >1,000 | ||||
| Northwest Woodland | ||||||||
| Oregon white oak-ponderosa pine | Replacement | 16% | 125 | 100 | 300 | |||
| Mixed | 2% | 900 | 50 | |||||
| Surface or low | 81% | 25 | 5 | 30 | ||||
| Ponderosa pine | Replacement | 5% | 200 | |||||
| Mixed | 17% | 60 | ||||||
| Surface or low | 78% | 13 | ||||||
| Oregon white oak | Replacement | 3% | 275 | |||||
| Mixed | 19% | 50 | ||||||
| Surface or low | 78% | 12.5 | ||||||
| Subalpine woodland | Replacement | 21% | 300 | 200 | 400 | |||
| Mixed | 79% | 80 | 35 | 120 | ||||
| Northwest Forested | ||||||||
| Sitka spruce-western hemlock | Replacement | 100% | 700 | 300 | >1,000 | |||
| Douglas-fir (Willamette Valley foothills) | Replacement | 18% | 150 | 100 | 400 | |||
| Mixed | 29% | 90 | 40 | 150 | ||||
| Surface or low | 53% | 50 | 20 | 80 | ||||
| Oregon coastal tanoak | Replacement | 10% | 250 | |||||
| Mixed | 90% | 28 | 15 | 40 | ||||
| Dry ponderosa pine (mesic) | Replacement | 5% | 125 | |||||
| Mixed | 13% | 50 | ||||||
| Surface or low | 82% | 8 | ||||||
| Douglas-fir-western hemlock (wet mesic) | Replacement | 71% | 400 | |||||
| Mixed | 29% | >1,000 | ||||||
| Mountain hemlock | Replacement | 93% | 750 | 500 | >1,000 | |||
| Mixed | 7% | >1,000 | ||||||
| Pacific silver fir (low elevation) | Replacement | 46% | 350 | 100 | 800 | |||
| Mixed | 54% | 300 | 100 | 400 | ||||
| Pacific silver fir (high elevation) | Replacement | 69% | 500 | |||||
| Mixed | 31% | >1,000 | ||||||
| Subalpine fir | Replacement | 81% | 185 | 150 | 300 | |||
| Mixed | 19% | 800 | 500 | >1,000 | ||||
| Great Lakes | ||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | ||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
|||||
| Great Lakes Grassland | ||||||||
| Mosaic of bluestem prairie and oak-hickory | Replacement | 79% | 5 | 1 | 8 | |||
| Mixed | 2% | 260 | ||||||
| Surface or low | 20% | 2 | 33 | |||||
| Great Lakes Woodland | ||||||||
| Northern oak savanna | Replacement | 4% | 110 | 50 | 500 | |||
| Mixed | 9% | 50 | 15 | 150 | ||||
| Surface or low | 87% | 5 | 1 | 20 | ||||
| Great Lakes Forested | ||||||||
| Northern hardwood maple-beech-eastern hemlock | Replacement | 60% | >1,000 | |||||
| Mixed | 40% | >1,000 | ||||||
| Conifer lowland (embedded in fire-resistant ecosystem) | Replacement | 36% | 540 | 220 | >1,000 | |||
| Mixed | 64% | 300 | ||||||
| Great Lakes floodplain forest | ||||||||
| Mixed | 7% | 833 | ||||||
| Surface or low | 93% | 61 | ||||||
| Great Lakes spruce-fir | Replacement | 100% | 85 | 50 | 200 | |||
| Minnesota spruce-fir (adjacent to Lake Superior and Drift and Lake Plain) | Replacement | 21% | 300 | |||||
| Surface or low | 79% | 80 | ||||||
| Maple-basswood | Replacement | 33% | >1,000 | |||||
| Surface or low | 67% | 500 | ||||||
| Maple-basswood mesic hardwood forest (Great Lakes) | Replacement | 100% | >1,000 | >1,000 | >1,000 | |||
| Maple-basswood-oak-aspen | Replacement | 4% | 769 | |||||
| Mixed | 7% | 476 | ||||||
| Surface or low | 89% | 35 | ||||||
| Northern hardwood-eastern hemlock forest (Great Lakes) | Replacement | 99% | >1,000 | |||||
| Oak-hickory | Replacement | 13% | 66 | 1 | ||||
| Mixed | 11% | 77 | 5 | |||||
| Surface or low | 76% | 11 | 2 | 25 | ||||
| Pine-oak | Replacement | 19% | 357 | |||||
| Surface or low | 81% | 85 | ||||||
| Eastern white pine-eastern hemlock | Replacement | 54% | 370 | |||||
| Mixed | 12% | >1,000 | ||||||
| Surface or low | 34% | 588 | ||||||
| Northeast | ||||||||
| Vegetation Community (Potential Natural Vegetation Group) | Fire severity* | Fire regime characteristics | ||||||
| Percent of fires | Mean interval (years) |
Minimum interval (years) |
Maximum interval (years) |
|||||
| Northeast Woodland | ||||||||
| Eastern woodland mosaic | Replacement | 2% | 200 | 100 | 300 | |||
| Mixed | 9% | 40 | 20 | 60 | ||||
| Surface or low | 89% | 4 | 1 | 7 | ||||
| Oak-pine (eastern dry-xeric) | Replacement | 4% | 185 | |||||
| Mixed | 7% | 110 | ||||||
| Surface or low | 90% | 8 | ||||||
| Northeast Forested | ||||||||
| Northern hardwoods (Northeast) | Replacement | 39% | >1,000 | |||||
| Mixed | 61% | 650 | ||||||
| Eastern white pine-northern hardwoods | Replacement | 72% | 475 | |||||
| Surface or low | 28% | >1,000 | ||||||
| Northern hardwoods-eastern hemlock | Replacement | 50% | >1,000 | |||||
| Surface or low | 50% | >1,000 | ||||||
| Northern hardwoods-spruce | Replacement | 100% | >1,000 | 400 | >1,000 | |||
| Appalachian oak forest (dry-mesic) | Replacement | 2% | 625 | 500 | >1,000 | |||
| Mixed | 6% | 250 | 200 | 500 | ||||
| Surface or low | 92% | 15 | 7 | 26 | ||||
| Beech-maple | Replacement | 100% | >1,000 | |||||
| Northeast spruce-fir forest | Replacement | 100% | 265 | 150 | 300 | |||
| *Fire Severities— Replacement: Any fire that causes greater than 75% top removal of a vegetation-fuel type, resulting in general replacement of existing vegetation; may or may not cause a lethal effect on the plants. Mixed: Any fire burning more than 5% of an area that does not qualify as a replacement, surface, or low-severity fire; includes mosaic and other fires that are intermediate in effects. Surface or low: Any fire that causes less than 25% upper layer replacement and/or removal in a vegetation-fuel class but burns 5% or more of the area [17,24]. |
||||||||
1. Andersen, Ulla Vogt; Calov, Birgitte. 1996. Long-term effects of sheep grazing on giant hogweed (Heracleum mantegazzianum). Hydrobiologia. 340(1-3): 277-284. [72376]
2. Buttenschon, Rita Merete; Nielsen, Charlotte. 2007. Control of Heracleum mantegazzianum by grazing. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 240-254. [73203]
3. Caffrey, J. M. 1999. Phenology and long-term control of Heracleum mantegazzianum. Hydrobiologia. 415: 223-228. [72322]
4. Caffrey, Joe M. 1994. Spread and management of Heracleum mantegazzianum along the Irish river corridors. In: de Waal, Louise C.; Child, Lois E.; Wade, P. Max; Brock, John H., eds. Ecology and management of invasive riverside plants. Biology Series. New York: John Wiley & Sons: 67-76. [72996]
5. Case, Martha A.; Beaman, John H. 1992. Heracleum mantegazzianum (giant cow parsnip): another exotic in the Michigan flora. Michigan Botanist. 31: 152-154. [72314]
6. Clegg, L. M.; Grace, J. 1974. The distribution of Heracleum mantegazzianum (Somm. & Levier) near Edinburgh. Transactions, Botanical Society of Edinburgh. 42: 223-229. [72975]
7. Czarapata, Elizabeth J. 2005. Invasive plants of the Upper Midwest: An illustrated guide to their identification and control. Madison, WI: The University of Wisconsin Press. 215 p. [71442]
8. Darbyshire, Stephen. 2004. Giant hogweed (Heracleum mantegazzianum) new to the Outaouais area in west Quebec. Trail and Landscape. 38(2): 75-79. [72315]
9. Dawe, Neil K.; White, Eric R. 1979. Giant cow parsnip (Heracleum mantegazzianum) on Vancouver Island, British Columbia. The Canadian Field Naturalist. 93(1): 82-83. [72379]
10. Dawson, F. Hugh; Holland, David. 1999. The distribution in bankside habitats of three alien invasive plants in the U.K. in relation to the development of control strategies. Hydrobiologia. 15: 193-201. [72328]
11. DiTomaso, Joseph M.; Healy, Evelyn A. 2003. Aquatic and riparian weeds of the West. Publication 3421. Davis, CA: University of California, Agriculture and Natural Resources. 442 p. [48834]
12. Dodd, Felicite S.; de Waal, Louise C.; Wade, P. Max; Tiley, Gordon E. D. 1994. Control and management of Heracleum mantegazzianum (giant hogweed). In: de Waal, Louise C.; Child, Lois E.; Wade, P. Max; Brock, John H., eds. Ecology and management of invasive riverside plants. New York: John Wiley & Sons: 111-126. [73015]
13. European and Mediterranean Plant Protection Organization. 2006. EPPO data sheet on invasive plants: Heracleum mantegazzianum, [Online]. In: EPPO activities on plant quarantine. Paris: European and Mediterranean Plant Protection Organization (Producer). Available: www.eppo.org/QUARANTINE/plants/Heracleum_mantgazzianum/HERMZ_ds.pdf [2009, April 8]. [72961]
14. Gleason, Henry A.; Cronquist, Arthur. 1991. Manual of vascular plants of northeastern United States and adjacent Canada. 2nd ed. New York: New York Botanical Garden. 910 p. [20329]
15. Grace, J.; Nelson, M. 1981. Insects and their pollen loads at a hybrid Heracleum site. New Phytologist. 87(2): 413-423. [72392]
16. Grime, J. P.; Mason, G.; Curtis, A. V.; Rodman, J.; Band, S. R.; Mowforth, M. A. G.; Neal, A. M.; Shaw, S. 1981. A comparative study of germination characteristics in a local flora. The Journal of Ecology. 69(3): 1017-1059. [70060]
17. Hann, Wendel; Havlina, Doug; Shlisky, Ayn; [and others]. 2008. Interagency fire regime condition class guidebook. Version 1.3, [Online]. In: Interagency fire regime condition class website. U.S. Department of Agriculture, Forest Service; U.S. Department of the Interior; The Nature Conservancy; Systems for Environmental Management (Producer). 119 p. Available: http://frames.nbii.gov/frcc/documents/FRCC_Guidebook_2008.07.10.pdf [2008, September 03]. [70966]
18. Huls, Jorg; Otte, Annette; Eckstein, R. Lutz. 2007. Population life-cycle and stand structure in dense and open stands of the introduced tall herb Heracleum mantegazzianum. Biological Invasions. 9(7): 799-811. [72330]
19. Jahodova, Sarka; Froberg, Lars; Pysek, Petr; Geltman, Dmitry; Trybush, Sviatlana; Karp, Angela. 2007. Taxonomy, identification, genetic relationships and distribution of large Heracleum species in Europe. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 1-19. [73193]
20. Kartesz, John T. 1999. A synonymized checklist and atlas with biological attributes for the vascular flora of the United States, Canada, and Greenland. 1st ed. In: Kartesz, John T.; Meacham, Christopher A. Synthesis of the North American flora (Windows Version 1.0), [CD-ROM]. Chapel Hill, NC: North Carolina Botanical Garden (Producer). In cooperation with: The Nature Conservancy; U.S. Department of Agriculture, Natural Resources Conservation Service; U.S. Department of the Interior, Fish and Wildlife Service. [36715]
21. Koutika, Lydie-Stella; Vanderhoeven, Sonia; Chapuis-Lardy, Lydie; Dassonville, Nicolas; Meerts, Pierre. 2007. Assessment of changes in soil organic matter after invasion by exotic plant species. Biology and Fertility of Soils. 44(2): 331-341. [72334]
22. Krinke, Lukas; Moravcova, Lenka; Pysek, Petr; Jarosik, Vojtech; Pergl, Jan; Perglova, Irena. 2005. Seed bank of an invasive alien, Heracleum mantegazzianum, and its seasonal dynamics. Seed Science Research. 15(3): 239-248. [72335]
23. Lamont, Eric E.; Young, Stephen M. 2004. Noteworthy plants reported from the Torrey Range--2002 and 2003. Journal of the Torrey Botanical Society. 131(4): 394-402. [72336]
24. LANDFIRE Rapid Assessment. 2005. Reference condition modeling manual (Version 2.1), [Online]. In: LANDFIRE. Cooperative Agreement 04-CA-11132543-189. Boulder, CO: The Nature Conservancy; U.S. Department of Agriculture, Forest Service; U.S. Department of the Interior (Producers). 72 p. Available: https://www.landfire.gov /downloadfile.php?file=RA_Modeling_Manual_v2_1.pdf [2007, May 24]. [66741]
25. LANDFIRE Rapid Assessment. 2007. Rapid assessment reference condition models, [Online]. In: LANDFIRE. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Lab; U.S. Geological Survey; The Nature Conservancy (Producers). Available: https://www.landfire.gov /models_EW.php [2008, April 18] [66533]
26. Massachusetts Invasive Plant Advisory Group (IMPAG). 2005. Strategic recommendations for managing invasive plants in Massachusetts. Final report: February 28, 2005, [Online]. Massachusetts Invasive Plant Advisory Group (Producer). Available: http://www.massaudubon.org/PDF/invasives/strategic_plan_042005.pdf [2009, January 30]. 15 p. [+ appendices]. [71599]
27. Mayer, LaRessa K. 2000. Comparison of management techniques for Heracleum mantegazzianum in north and central Europe. Restoration and Reclamation Review--Student Online Journal. Volume 6--Fall. St. Paul, MN: University of Minnesota, Department of Horticultural Science. Available: http://horticulture.cfans.umn.edu/vd/h5015/00papers/mayer.htm [2009, March 27]. [72316]
28. Mehrhoff, L. J.; Silander, J. A., Jr.; Leicht, S. A.; Mosher, E. S.; Tabak, N. M. 2003. Have you seen this plant? Giant hogweed (Heracleum mantegazzianum), [Online]. In: Noxious weeds. In: IPANE: Invasive Plant Atlas of New England. Storrs, CT: University of Connecticut, Department of Ecology and Evolutionary Biology (Producer). Available: http://nbii-nin.ciesin.columbia.edu/ipane/ipanespecies/HogweedWarning.pdf [2009, March 31]. [73155]
29. Moravcova, Lenka; Perglova, Irena; Pysek, Petr; Jarosik, Vojtech; Pergl, Jan. 2005. Effects of fruit position on fruit mass and seed germination in the alien species Heracleum mantegazzianum (Apiaceae) and the implications for its invasion. Acta-Oecologica. 28(1): 1-10. [72338]
30. Moravcova, Lenka; Pysek, Petr; Krinke, Lukas; Pergl, Jan; Perglova, Irena; Thompson, Ken. 2007. Seed germination, dispersal, and seed bank in Heracleum mantegazzianum. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 74-91. [73196]
31. Moravcova, Lenka; Pysek, Petr; Pergl, Jan; Perglova, Irena; Jarosik, Vojtech. 2006. Seasonal pattern of germination and seed longevity in the invasive species Heracleum mantegazzianum. Preslia. 78(3): 287-301. [72386]
32. Morton, J. K. 1975. The giant cow-parsnip, Heracleum mantegazzianum Umbelliferae, in Canada. The Canadian Field Naturalist. 89: 183-184. [72318]
33. Morton, J. K. 1978. Distribution of giant cow-parsnip (Heracleum mantegazzianum) in Canada. The Canadian Field Naturalist. 92: 182-185. [72319]
34. Mullerova, Jana; Pysek, Petr; Jarosik, Vojtech; Pergl, Jan. 2005. Aerial photographs as a tool for assessing the regional dynamics of the invasive plant species Heracleum mantegazzianum. Journal of Applied Ecology. 42(6): 1042-1053. [72339]
35. New Jersey Department of Agriculture. 2005. Giant hogweed (Heracleum mantegazzianum): An attractive but dangerous noxious weed--Have you seen this plant? Trenton NJ: New Jersey Department of Agriculture. Pamphlet. [72411]
36. New York State Department of Agriculture and Markets. [2005]. Giant hogweed (Heracleum mantegazzianum). An attractive but dangerous noxious weed. Have you seen this plant? Albany, NY: New York State Department of Agriculture and Markets, Division of Plant Industry; U.S. Department of Agriculture, Animal & Plant Health Inspection Service, Plant Protection & Quarantine. 4 p. [72960]
37. Nielsen, Charlotte; Hartvig, Per; Kollmann, Johannes. 2008. Predicting the distribution of the invasive alien Heracleum mantegazzianum at two different spatial scales. Diversity and Distributions. 14(2): 307-317. [72343]
38. Nielsen, Charlotte; Heimes, Christine; Kollmann, Johannes. 2008. Little evidence for negative effects of an invasive alien plant on pollinator services. Biological Invasions. 10(8): 1353-1363. [72344]
39. Nielsen, Charlotte; Vanaga, Ineta; Treikale, Olga; Priekule, Ilze. 2007. Mechanical and chemical control of Heracleum mantegazzianum and H. sosnowskyi. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 226-239. [73202]
40. Oregon Department of Agriculture. 2008. Toxic plant alert!: Giant hogweed--Heracleum mantegazzianum. Salem OR: Oregon Department of Agriculture, Plant Division, Noxious Weed Control Program. Pamphlet. [72412]
41. Otte, Annette; Franke, Rene. 1998. The ecology of the Caucasian herbaceous perennial Heracleum mantegazzianum Somm. et Lev. (giant hogweed) in cultural ecosystems of Central Europe. Phytocoenologia. 28(2): 205-232. [72345]
42. Page, Nicholas A.; Wall, Ronald E.; Darbyshire, Stephen J.; Mulligan, Gerald A. 2006. The biology of invasive alien plants in Canada. 4. Heracleum mantegazzianum Sommier & Levier. Canadian Journal of Plant Science. 86(2): 569-589. [72346]
43. Parks, Catherine G.; Radosevich, Steven R.; Endress, Bryan A.; Naylor, Bridgett J.; Anzinger, Dawn; Rew, Lisa J.; Maxwell, Bruce D.; Dwire, Kathleen A. 2005. Natural and land-use history of the Northwest mountain ecoregions (USA) in relation to patterns of plant invasions. Perspectives in Plant Ecology, Evolution and Systematics. 7: 137-158. [70353]
44. Pergl, Jan; Huls, Jorg; Perglova, Irena; Eckstein, R. Lutz; Pysek, Petr; Otte, Annette. 2007. Population dynamics of Heracleum mantegazzianum. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 92-111. [73197]
45. Pergl, Jan; Perglova, Irena; Pysek, Petr; Dietz, Hansjorg. 2006. Population age structure and reproductive behavior of the monocarpic perennial Heracleum mantegazzianum (Apiaceae) in its native and invaded distribution ranges. American Journal of Botany. 93(7): 1018-1028. [72347]
46. Perglova, Irena; Pergl, Jan; Pysek, Petr. 2006. Flowering phenology and reproductive effort of the invasive alien plant Heracleum mantegazzianum. Preslia. 78(3): 265-285. [72348]
47. Perglova, Irena; Pergl, Jan; Pysek, Petr. 2007. Reproductive ecology of Heracleum mantegazzianum. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 55-73. [73195]
48. Pojar, Jim; MacKinnon, Andy, eds. 1994. Plants of the Pacific Northwest coast: Washington, Oregon, British Columbia and Alaska. Redmond, WA: Lone Pine Publishing. 526 p. [25159]
49. Pysek, Petr. 1991. Heracleum mantegazzianum in the Czech Republic: dynamics of spreading from the historical perspective. Folia Geobotanica et Phytotaxonomica. 26(4): 439-459. [72351]
50. Pysek, Petr; Jarosik, Vojtech; Mullerova; Jana; Pergl, Jan; Wild, Jan. 2008. Comparing the rate of invasion by Heracleum mantegazzianum at continental, regional, and local scales. Diversity and Distributions. 14(2): 355-363. [72349]
51. Pysek, Petr; Kopecky, Miroslav; Jarosik, Vojtech; Kotkova, Pavla. 1998. The role of human density and climate in the spread of Heracleum mantegazzianum in the Central European landscape. Diversity and Distributions. 4(1): 9-16. [72391]
52. Pysek, Petr; Krinke, Lukas; Jarosik, Vojtech; Perglova, Irena; Pergl, Jan; Moravcova, Lenka. 2007. Timing and extent of tissue removal affect reproduction characteristics of an invasive species Heracleum mantegazzianum. Biological Invasions. 9(3): 335-351. [72350]
53. Pysek, Petr; Morvcova, L. 2009. [Email to Corey L. Gucker]. March 6. Heracleum mantegazzianum: Recent seed bank findings. Pruhonice, Czech Republic: Academy of Sciences of the Czech Republic, Institute of Botany. On file with: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; RWU 4403 files. [73479]
54. Pysek, Petr; Mullerova, Jana; Jarosik, Vojtech. 2007. Historical dynamics of Heracleum mantegazzianum invasion at regional and local scales. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 42-54. [73194]
55. Pysek, Petr; Perglova, Irena; Krinke, Lukas; Jarosik, Vojtech; Pergl, Jan; Moravcova; Lenka. 2007. Regeneration ability of Heracleum mantegazzianum and implications for control. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 112-125. [73198]
56. Pysek, Petr; Prach, Karel. 1993. Plant invasions and the role of riparian habitats: comparison of four species alien to central Europe. Journal of Biogeography. 20(4): 413-420. [72354]
57. Pysek, Petr; Pysek, Antonin. 1995. Invasion by Heracleum mantegazzianum in different habitats in the Czech Republic. Journal of Vegetation Science. 6(5): 711-718. [72355]
58. Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford: Clarendon Press. 632 p. [2843]
59. Raven, Hans Peter; Treikale, Olga; Vanaga, Ineta; Priekule, Ilze. 2007. Revegetation as a part of an integrated management strategy for large Heracleum species. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 272-283. [73225]
60. Roche, Cindy. 1992. Weeds--Giant hogweed (Heracleum mantegazzianum Somm. & Lev.). PNW Bulletin 429. Corvallis, OR: Pacific Northwest Extension Service. 2 p. [72360]
61. Sheppard, A. W.; Shaw, R. H.; Sforza, R. 2006. Top 20 environmental weeds for classical biological control in Europe: a review of opportunities, regulations and other barriers to adoption. Weed Research. 46(2): 93-117. [72388]
62. Smither-Kopperl, Margaret. 2007. The first line of defense: interceptions of federal noxious weed seeds in Washington. In: Harrington, Timothy B.; Reichard, Sarah H., tech. eds. Meeting the challenge: invasive plants in Pacific Northwest ecosystems. Gen. Tech. Rep. PNW-GTR-694. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 19-22. [69617]
63. Stewart, F.; Grace, J. 1984. An experimental study of hybridization between Heracleum mantegazzianum Somm. & Levier and Heracleum sphondylium L. subsp. sphondylium (Umbelliferae). Watsonia. 15(2): 73-83. [72362]
64. Stickney, Peter F. 1989. Seral origin of species comprising secondary plant succession in Northern Rocky Mountain forests. FEIS workshop: Postfire regeneration. Unpublished draft on file at: U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Fire Sciences Laboratory, Missoula, MT. 10 p. [20090]
65. Theile, Jan; Otte, Annette. 2006. Analysis of habitats and communities invaded by Heracleum mantegazzianum Somm. et Lev. (giant hogweed) in Germany. Phytocoenologia. 36(2): 281-320. [72365]
66. Theile, Jan; Otte, Annette. 2007. Impact of Heracleum mantegazzianum on invaded vegetation and human activities. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 144-156. [73200]
67. Theile, Jan; Otte, Annette. 2008. Invasion patterns of Heracleum mantegazzianum in Germany on the regional and landscape scales. Journal for Nature Conservation. 16(2): 61-71. [72389]
68. Theile, Jan; Otte, Annette; Eckstein, R. Lutz. 2007. Ecological needs, habitat preferences and plant communities invaded by Heracleum mantegazzianum. In: Pysek, P.; Cock, M. J. W.; Nentwig, W.; Ravn, H. P., eds. Ecology and management of giant hogweed (Heracleum mantegazzianum). Wallingford, UK; Cambridge, MA: CAB International: 126-143. [73199]
69. Theile, Jan; Schuckert, Ulrike; Otte, Annette. 2008. Cultural landscapes of Germany are patch-corridor-matrix mosaics for an invasive megaforb. Lanscape Ecology. 23(4): 453-465. [72363]
70. Tiley, G. E. D.; Dodd, Felicite S.; Wade, P. M. 1996. Biological flora of the British Isles. Heracleum mantegazzianum Sommier & Levier. Journal of Ecology. 84(2): 297-319. [72366]
71. Tiley, Gordon E. D.; Philip, Bruce. 1997. Observations on flowering and seed production in Heracleum mantegazzianum in relation to control. In: Brock, J. H.; Wade, M; Pysek, P.; Green, D., eds. Plant invasions: studies from North America and Europe. Leiden, The Netherlands: Backhuys Publishers: 123-137. [73561]
72. U.S. Department of Agriculture, Animal and Plant Health Inspection Service (APHIS). 2006. Federal noxious weed list, [Online]. In: Plant health--Noxious weeds program. Washington, DC: U.S. Department of Agriculture, Animal and Plant Health Inspection Service (Producer). Available: http://www.aphis.usda.gov/plant_health/plant_pest_info/weeds/downloads/weedlist2006.pdf [2009, April 8]. [36689]
73. U.S. Department of Agriculture, Natural Resources Conservation Service. 2009. PLANTS Database, [Online]. Available: https://plants.usda.gov /. [34262]
74. Voss, Edward G. 1985. Michigan flora. Part II. Dicots (Saururaceae--Cornaceae). Bull. 59. Bloomfield Hills, MI: Cranbrook Institute of Science; Ann Arbor, MI: University of Michigan Herbarium. 724 p. [11472]
75. Washington State Noxious Weed Control Board. 2007. Information about giant hogweed (Heracleum mantegazzianum), [Online]. In: Written findings of the State Noxious Weed Control Board--Class A weed. Washington State Noxious Weed Control Board (Producer). Available: http://www.nwcb.wa.gov/weed_info/written_findings/Heracleum_mantegazzianum.html [ 2009, March 27]. [73017]
76. Westbrooks, Randy G. 1998. Invasive plants: changing the landscape of America. Fact Book. Washington, DC: Federal Interagency Committee for the Management of Noxious and Exotic Weeds. 109 p. [33874]
77. Willis, S. G.; Hulme, P. E. 2002. Does temperature limit the invasion of Impatiens glandulifera and Heracleum mantegazzianum in the UK? Functional Ecology. 16(4): 530-539. [72374]
78. Willoughby, Ian. 1996. Noxious weeds. Research Information Note 274. Edinburgh, UK: Forestry Commission, Research Division. 8 p. [72375]