| FEIS Home Page |
 |
| Figure 1—Desert-willow in Anza Borrego State Park, California. Photo © 2000 Larry Blakely, used with permission. |
| This review summarizes the information on the biology, ecology, and effects of fire on desert-willow in that was available in the scientific literature as of 2021.
Desert-willow is native to the southwestern United States and northern Mexico, occurring in the Mojave, Sonoran, Colorado, southern Great Basin, and Chihuahuan deserts. It is an obligate riparian species, and it is most common in wash shrublands and mixed-deciduous riparian woodlands. Desert-willow reproduces sexually and regenerates from seed. It sprouts from the root crown following top-kill. Desert-willow is cross-pollinated by bumble bees. Seed crops are typically large in wet years. Germination occurs on moist substrates, soon after flooding, and growth is typically rapid in moist soils. Desert-willow is most common in open areas. Flooding is the primary disturbance in communities with desert-willow, but desert-willow also grows in early postfire environments. With alterations in hydrology and climate change, fire is becoming the primary disturbance in many riparian ecosystems. Low- to moderate-severity fire prunes or top-kills desert-willow. It sprouts after top-kill by fire and may also establish from seed. Historically, fuels in plant communities where desert-willow is common were likely mostly sparse and discontinuous, although information on this is limited. Fires were probably infrequent, and they likely varied in severity and extent across southwestern riparian ecosystems. Most fires were likely patchy and of low severity due to patchy and sparse fuels. Changes in hydrology and vegetation have undoubtedly affected fuels and fire regimes of southwestern riparian communities and surrounding uplands. However, the degree to which fire regimes are changing in southwestern riparian ecosystems—or what the effects might be for riparian ecosystem structure, function, and resilience—are not well known. Severe, intense, and/or frequent fire can interact with altered hydrology to accelerate the replacement of native plants with nonnative and often invasive plants such as tamarisk and Russian-olive. When nonnative species invade the overstory and/or understory and alter riparian vegetation composition and structure, fuel characteristics may also change. In turn, this can alter fire behavior and fire regimes. Fire management in southwestern riparian areas may include fuel reduction and prescribed fire treatments that promote or maintain native vegetation and reduce or eradicate invasives. The value of southwestern riparian woodlands for both biotic communities and human livelihoods is well established. Communities with desert-willow provide important habitat and food for a wide variety of wildlife species. Riparian areas with desert-willow provide many ecosystem services including recreational opportunities, harboring pollinators, and providing livestock forage. Few naturally functioning riparian areas remain in the Southwest, and those that do are threatened with climate change. Southwestern riparian ecosystems are vulnerable to decline under increasingly hotter and drier conditions. Climate change is expected to alter water flow and lower water tables, and to increase the frequency and severity of extreme weather. This will likely result in more frequent and severe floods as well as more intense droughts. Where natural flow regimes are more intact and tamarisk and other invasive species are suppressed by native vegetation, desert-willow and other native trees are more likely to maintain dominance in postfire landscapes. |
Citation:
Fryer, Janet L. 2022. Chilopsis linearis, desert-willow.
In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service,
Rocky Mountain Research Station, Missoula Fire Sciences Laboratory (Producer).
Available: www.fs.usda.gov/database/feis/plants/tree/chilin/all.html
[].
Chilopsis linearis subsp. arcuate (Fosberg) Henderson
Chilopsis linearis subsp. linearis [6,85,97,119,161,220,224]
Chilopsis linearis subsp. tomenticaulis Henrickson [85]
Common names are used throughout this Species Review. For scientific names of plant and animal species mentioned in this review and links to other FEIS Species Reviews, see the Appendix.
Reviews cited in this Species Review include: [127,134,192,220,244].
SYNONYMS
For Chilopsis linearis subsp. arcuate:
Chilopsis linearis (Cav.) Sweet var. arcuate Fosberg [68]
For Chilopsis linearis subsp. linearis:
Chilopsis linearis (Cav.) Sweet var. glutinosa [1,68,100,129]
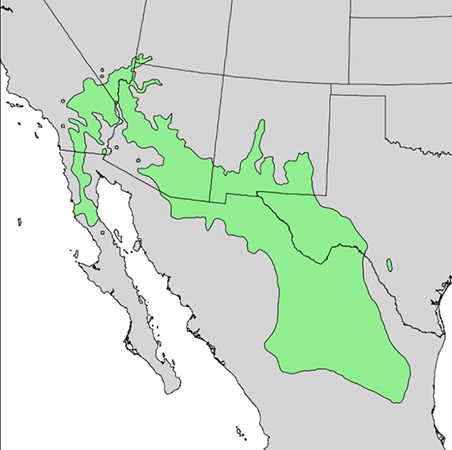 |
| Figure 2—Distribution of desert-willow. Map from Little (1976) [120] and digitized by Thompson et al. (1999) [213]. Desert-willow also occurs in Kansas, Oklahoma, and Georgia, although precise distributional information is sparse for those states [96,224]. |
Desert-willow is native to the southwestern United States and northern Mexico [96,120,224] (fig. 2), occurring in the Mojave, Sonoran, Colorado, southern Great Basin, and Chihuahuan deserts [51,159]. It is distributed from southern California eastward through southern Utah to western Texas [63,96,100,120,127,130,226] and southward to Baja California Norte, Durango, and Zacatecas, Mexico [120,130]. Disjunct but apparently native populations occur in Kansas, Oklahoma, and Georgia [96,224]. Desert-willow is planted outside its native range [63].
Chilopsis linearis subsp. arcuate occurs in California and Arizona; Chilopsis linearis subsp. linearis occurs in Kansas, Oklahoma, and Texas. Distributions of these subspecies overlap in Socorro County, New Mexico [224]. Chilopsis linearis subsp. tomenticaulis occurs in northeastern Mexico [85]. PLANTS Database provides distributional maps for the first two subspecies.
States:
United States: AZ, CA, GA, KS, NV, NM, OK, TX, UT [96,120,224]
Mexico: BCN, Chih, Coah, Dgo, Son, Zac [120]
SITE CHARACTERISTICS
Desert-willow is a warm-desert species that grows in lowlands [220,244] with bimodal rainfall. In its northwestern distribution, rain falls mostly in winter; in its eastern distribution, rain falls mostly in summer [9,156,183,220,236].
Arid climate restricts this phreatophyte to moist areas [51,159]; desert-willow is an obligate outwash or riparian species [13,19,39,53,54,102,157]. It is considered an indicator of sites with relatively shallow water tables [138]. Sites with desert-willow include stream- and riverbanks, alluvial fans, washes, arroyos, gullies [10,73,116,220,243,244], and bajadas [51,159,215,251]. It also grows on moist plains, hillsides [51,159,251], mesas, and terraces [51,159,215,226,251]. It is rare on unstable dunes [220].
Desert-willow is flood tolerant [16,73,92,156], and it is frequent on sites that experience intermittent flooding [10,73,116,146,159,220,243,244]. It often occupies the middle and banks of drainage channels, sometimes covering broad expanses in washes [73,92]. On the Gila River Resource Area, Arizona, desert-willow is most frequent in areas that receive severe scouring [137]. It may survive 1 to 3 months of inundation during the growing season, going dormant during prolonged floods [16]. It is somewhat drought intolerant ([210,217]); desert-willow mainly avoids drought by reducing transpiration [149,150] and/or shedding its leaves (see Seasonal Development).
Desert-willow grows at low elevations, occurring between 450 and 1,830 m elevation across its range [98,100,127], usually below 1,600 m [51,159,215] (table 1).
| Table 1—Elevational range of desert-willow by area. | |
Area |
Elevational range (m) |
Sonoran Desert |
≤1,500 [220] |
| Southwest | 1,146–1,433 [208] |
| Arizona | generally ≤1,650 [166], rarely to 1,830 [100] |
| California | <2,100 [6] |
| New Mexico | 1,200–1,700 [34,129] |
| Nevada, southern | 500–1,800 [98] |
| Texas, Trans-Pecos region | 600–1,500 [161] |
| Utah, southern | 1,400 [91] |
Desert-willow generally grows in alluvial substrates [73,239], with soils composed of sandy loam [95], sand [6,34,38,141,154,157,239,246], gravel [6,46,129,130,157,226,235,251], and/or cobble [157,159]. Desert-willow is intolerant of saline soils [182].
PLANT COMMUNITIES
Desert-willow is dominant in or a common component of wash shrublands [19,21,112,125,162,166,176,214,215,239]; mixed-deciduous riparian [12,56,84,165], pinyon-oak [115,134,147], and Joshua tree [43] woodlands; and some grasslands [76,238]. Washes and woodlands with desert-willow are typically adjacent to upland desert shrubland, desert grassland, or plains grassland communities [19,159,183,211].
Desert-willow is most common in and may codominate in creosote bush [43,113,184], mesquite [20], and catclaw acacia [21,214,215,242,247] wash shrublands. In southern California and Nevada, desert-willow grows in creosote bush-burrobush [43,113,184] and blackbrush [159,183] communities. In Arizona, it commonly grows in washes with catclaw acacia, desertbroom, eastern Mojave buckwheat, screwbean mesquite, and/or velvet mesquite [21,154,159,242]; and in riparian bosques and scrublands with honey mesquite [21,154,215,247]. In the Colorado, Sonoran, and Chihuahuan deserts, desert-willow sometimes codominates or associates with paloverde [21,215,247]. In Guadalupe Mountains National Park, western Texas [75], and southward to northern Mexico [42,64], it codominates with creosote bush and honey mesquite [42,64,75].
Desert-willow also occurs in other wash shrublands, or other shrublands over shallow water tables. In the Coachella Valley, California, smoketree-desert-willow-burrobrush communities occur near drier yucca/cactus communities [246]. Desert-willow is locally dominant in paloverde-cactus communities of the Mojave and Sonoran deserts [139,159,239]. In Arizona, it may also codominate with spiny hackberry and western soapberry [148], and on the Kofa National Wildlife Refuge, Arizona, desert-willow grows with ironwood, blue paloverde, and smoketree [245]. In the Trans-Pecos region of Texas, desert-willow is a component of Apache plume shrublands along arroyos [211]. In Chihuahua, Mexico, desert-willow grows in tropical-arid riparian shrublands with baccharis, singlewhorl burrobrush, and tree tobacco [156].
Desert-willow is a subcanopy tree in many riparian mixed-deciduous woodlands of the Southwest. These woodlands are codominated by Arizona sycamore, Arizona walnut, Fremont cottonwood, and/or velvet ash [12,84,165]. Willows—including arroyo willow, Goodding’s willow, and narrowleaf willow [12,155]—often codominate as well [56,155]. Shorter-statured deciduous communities dominated by desert-willow mostly occur in lower portions of arroyos, where streambeds widen [159]. Desert-willow riparian woodlands are a minor type in the Santa Ana Mountains of California [233], central New Mexico [56], and western Texas [159]. In southern Utah, desert-willow forms thickets with cottonwoods and nonnative tamarisk [98]. On the Stockton Plateau of west-central Texas, Arizona walnut-desert-willow communities occur on streambanks [86]. Desert-willow/splitleaf brickellbush communities occur on the Edwards Plateau of south-central Texas, particularly on intermittently flooded banks and washes [144]. Little walnut-desert-willow-splitleaf brickellbush communities occur in southern Texas [145]. Along the Rio Conchos in Chihuahua, Mexico, desert-willow is subdominant in black walnut-Lombardy poplar-Texas paloverde communities [42].
Desert-willow occurs in moist areas within pinyon-juniper-oak woodlands [115,134,147]. At the turn of the last century, Leiberg (1900) noted Fremont cottonwood-velvet ash/desert-willow-white alder stands in ravines within singleleaf pinyon-Parry pinyon-canyon live oak woodlands of the San Jacinto Reserve in California (now part of the San Bernardino National Forest) [115]. In the southern Guadalupe Mountains of Texas, desert-willow is a component of wavyleaf oak-alligator juniper-Texas madrone woodlands [147].
Desert-willow occurs adjacent to and may finger into some desert [167,230] and southern plains [76,211,238] grasslands. It grows in upper-elevation (1,700-1,900 m) [238] blue grama-tobosagrass [76,238] desert or plains grasslands. In Mexico, desert-willow codominates or occurs in desertbroom-netleaf hackberry-spiny hackberry riparian scrub [167] that merges into desert grasslands [230] composed of Arizona muhly, Hall's panicgrass, and sideoats grama [167].
See table A3 for a list of representative plant community classifications in which desert-willow occurs.BOTANICAL DESCRIPTION
This description covers characteristics that may be relevant to fire ecology and is not meant for identification. Identification keys are available (e.g., [1,6,43,55,129,251]). Henrickson (1985) provides highly detailed botanical descriptions of desert-willow and its subspecies [85].
Aboveground Description
Desert-willow is a deciduous [85,92,127,130,232] small tree or shrub [1,127,129]. It is typically 3 to 7.5 m tall [127] but sometimes up to 12 m tall [1]. Habit is described as "tall and straggling" [180]
(fig. 1).
Trunks range from 10 to 40 cm in diameter [1], with one to several stems arising from the root crown [91,129,251]. The bark is "very thin": not more than 0.6 cm thick [91]. The upper crown is spreading [119]. Like its namesake, willow, desert-willow has slender, flexible branchlets [68] and drooping, elongated (linear-lanceolate) leaves [46,68,85,155,237]. Leaves are simple [6,100,130,251], with a thick, waxy cuticle that reduces water loss [220,226]. Some leaves may have extrafloral nectaries [85,157]. In a Sonoran Desert population, presence of extrafloral nectaries was variable both within and among individual trees [157]. The inflorescence is a raceme or panicle with showy, tubular flowers [85,129,161,251]
(fig. 3).
The fruit is a slender capsule [127,129,251] from 10 to 30 cm long and about 6 mm in diameter [119,127,237]. In part, this genus/species is distinguished by its long, leathery capsules. Its seeds have a fringe of soft white hairs on each end [85,127,130,161,180,232,251]
(fig. 4).
Seeds are flat, about 8 mm long [127], and have 10 to 15 mm-long hairs [129,161,232,251].
Belowground Description
Desert-willow has a taproot [46] and is deep rooted [51,116,180,226]. Horizontal roots may extend as far as 15 m [220]. On the Jornada Experimental Range, New Mexico, the taproot of a 50-m³ tree was 160 cm deep. Most of the total root biomass was in shallow soil: the distribution of root biomass by soil depth averaged 72% from 0 to 40 cm, 18% from 40 to 80 cm, 8.6% from 80 to 120- cm, and 1.4% from 140 to 160 cm [51].
Stand or Population Structure
Structure of plant communities with desert-willow are variable in structure [175]. Desert-willow is typically a subcanopy tree in mixed-deciduous communities, but it is a canopy species in shrublands or short-statured woodlands. It often forms thickets along washes [91]. Communities in scour zones [154] or that are accessible to off-highway vehicles (OHVs) [69] may lack or have sparse understories.
A few studies quantify stand structure of desert-willow in pure and mixed-deciduous communities. See these references for densities of desert-willow and associated trees in Arizona and New Mexico [208], New Mexico [51], and northwestern Chihuahua, Mexico [208].
Desert-willow may live as long as 100 years [55]. Other information on age class structure was lacking in the literature.
Raunkiaer Life Form
Phanerophyte
Geophyte [164]
SEASONAL DEVELOPMENT
Desert-willow has a long flowering period (table 2). It may produce flowers from April through August throughout its range [127]. Flowering is triggered by winter [220] and summer [138,220] rains. Mass flowering usually occurs early in the growing season. After mass flowering, plants usually produce only one to four flowers at a time, over an extended flowering season. Flowers open and produce nectar from late afternoon through late night, with flowers senescing after 1 to 2 days [172]. Fruits ripen from late summer to late fall and persist on the plant through winter [127]. Seed dispersal coincides with winter and spring floods [101]. Seeds germinate in summer in southern California [246]; other germination times were not available.
Desert-willow may drop its leaves in any season. It is cold-deciduous [46,102,116,220], with leaves falling in late fall or winter [46,116] after temperatures fall below around 5 °C [220]. It is also drought-deciduous [149,155,177,220].
| Table 2—Phenology of desert-willow. | |
| Area | Event |
| Intermountain region | Flowers May-July [43] |
| Sonoran Desert | Leaves 1st emerge March-May; flowers April-August [172,220]; drought-induced leaf drop June-July; 2nd leaf emergence with August rains [220] |
| Southwest | Flowers May-June; sporadically thereafter following summer rains [232] |
| Arizona | Flowers April-August [100] |
| California | Germinates in summer [246]; flowers May-June [6,28] |
| New Mexico | Flowers April-August [129,237]; leaf drop October-November [102] |
| Nevada, southern | Flowers May-August [98] |
| Texas, Trans-Pecos region | Flowers April-September [161] |
| Texas, north-central | Flowers May-September [55] |
| Utah, Washington County | Flowers May-July [226] |
| Baja California | Flowers August-April [251] |
Pollination and Breeding System
The flowers are perfect [127], with a strong, pleasant scent [130,188,226]. Sonoran bumble bees [43,171,172,226,248] are the main pollinators. Hummingbirds [22,80,171,172,220,226], honeybees [171,172,232], lady-beetles, small flies [220], hawkmoths, carpenter bees [43,171,172,226], and other nectar-robber bees [171,172] also visit the flowers; but they are not effective pollinators [171,172].
Desert-willow is mostly cross-pollinated [158,172]. Its flowers close after the stigmata are touched, reopening if pollination was ineffective (e.g., not enough pollen was deposited or self-pollination occurred) [172]. Hand-pollination of flowers near Tucson, Arizona, resulted in a 540% increase in fruit set compared to bumble bee pollination [158].
Seed Production
Desert-willow typically produces good seed crops, although quantitative data are lacking for its seed production. Numerous flowers are produced early in the growing season
(see Seasonal Development)
[158], when a single tree may produce hundreds of flowers [172]. Subsequently, numerous seeds are contained in each capsule [43,161]. Flower and fruit production are positively associated with precipitation; plants may not produce flowers in a very dry year [51]
(see Plant Growth). Fruit set may be limited by insufficient amounts of outcrossed pollen (e.g., inadequate movement of pollinators among individual plants) [158].
Seed Dispersal
The seeds are dispersed by wind [14] and water [101]. The seed hairs
(fig. 4) aid dispersal [14].
Seed Banking
Desert-willow has a transient [122,127], soil-stored seed bank [50,127]. Viable desert-willow seed was collected from soil in southern Nevada for the Millennial Seed Bank Project [50], although details of collection were not provided.
Germination and Seedling Establishment
Desert-willow seeds are not dormant [127,128,180,254] and
nonrefractory
(Fryer 2022, this publication). They probably remain viable only until late in the spring following dispersal [128]. Fresh seeds germinate “readily” [220], although a short period of
afterripening
enhances germination [127]. Germination rates are generally high, ranging from about 50% [232] to 80% [127]. In a laboratory test, fresh seeds averaged 70% germination in the dark, with germination reduced to 15% for seeds dry-stored for 6 months [122].
A moist substrate is required for germination, and flooding and light shade promote germination and recruitment of desert-willow [141,191,200,243]. During floods, desert-willow seeds often lodge under boulders, which provide shade for emergents [38]. Desert-willow may also establish near its own canopy [141].
Plant Growth
Seedling growth is rapid under favorable conditions [114,126,180]. Desert-willow seedlings may grow as much as 1 m/year if water is not limiting [220]. In the greenhouse, desert-willow seedlings averaged 20.6 cm tall 20 days after emerging, and roots averaged 15.0 cm long [180]. In Lubbock, Texas, desert-willow seedlings planted in gravel averaged 815 g dry weight at initial planting and 1,093 g dry weight at 1 year old. Height gain after 1 year averaged 106 cm [105].
Desert-willow's rate of photosynthesis is relatively high [199], suggesting that it maintains relatively rapid growth rates. A study near Las Cruces, New Mexico, showed desert-willow maintains relatively constant photosynthetic rates and water potentials despite either drought or rapid fluctuations in water availability [46].
Growth rate in a wet year may be double that in a dry year. On an arroyo on the New Mexico State University's Agricultural Experiment Station, the mean number of new desert-willow shoots/plant was 440 over 4 years, with a range from 72 to 960 shoots/plant. The rate and amount of growth was strongly associated with available moisture, based on precipitation. Lowest production of leaf biomass (3 kg/ha) occurred in a year (1971) with a dry spring and relatively low summer precipitation. Almost no fruits were produced that year. The next growing season (1972–1973) had relatively continuous moisture from fall through summer. This resulted in the largest peak of leaf (9 kg/ha) and fruit (2.5 kg/ha) biomass production in the 4 years of study [51].
Dams limit the extent of flooding and germination of riparian trees within the floodplain, and they lower the water table in areas away from the active channel. Thus, seedlings and established trees are more likely to die. Changes to peak discharge timing also limit reproduction of riparian species [192].
Vegetative Reproduction and Regeneration
Desert-willow sprouts from the root crown [18,218,220] after
top-kill by fire
[18,218], flood damage [18], or cutting [119]. It may also
layer [57].
Mortality
Drought causes mortality of riparian species such as desert-willow, which depend on floods and groundwater for survival and growth [192]. Seedling and sprout mortality may be high during hot, dry periods and/or when water tables drop [243]. No further information was available on mortality of desert-willow except in the context of
fire.
SUCCESSIONAL STATUS
Desert-willow is most common in open areas [137], although it is somewhat shade tolerant and can establish near parent plants [141].
Flooding is the primary disturbance in mixed-deciduous riparian communities of the Southwest [67,191,200,243]. Due to erosion and deposition during floods, these communities often experience large-scale disturbances that set back succession [30,134,243]. Early-successional stages are common in mixed-deciduous woodlands [30,243], and trees on heavily scoured sites may never reach maturity [243] (i.e., the stem exclusion stage). In the Sonoran Desert in western Chihuahua, Mexico, sporadic floods on the lower banks of major rivers with scrubby desert-willows are described as "torrential" [156]. Trees on upper banks of such riparian areas are most likely mature [243], while trees on lower banks may be dislodged in floods.
Desert-willow also establishes in early postfire environments (see Fire Adaptations and Plant Response to Fire). Fire is becoming the primary disturbance in riparian ecosystems due to changes in hydrology (see Fire Management Considerations), climate, and plant community composition and structure (i.e., fuels) [245] (also see Fire Regimes).
Because it is unpalatable and associated species are preferred, desert-willow cover may increase with overgrazing [64,138]. In Coahuila, Mexico, the importance value of desert-willow was higher on sites with livestock (cattle, domestic goats, donkeys, and horses) than on sites without livestock [64]. However, near Little Ash Creek on the Prescott National Forest, Arizona, desert-willow was present only in livestock exclosures [209], suggesting that trampling and/or browsing of desert-willow had occurred in unprotected areas.IMMEDIATE FIRE EFFECTS
Low- to moderate-severity fire prunes or top-kills desert-willow [12,18,218,219,244]. Its "very thin" bark [91] provides little protection from fire. Although desert-willow typically sprouts after top-kill by fire (see below), it may not survive severe or repeated fire [12,244].
Postfire Regeneration Strategy
Tree with a sprouting root crown
Tall shrub with a sprouting root crown [198]
Other Possible Strategies:
Ground residual colonizer (on site, initial community)
Initial off-site colonizer (off site, initial community)
Secondary colonizer (on- or off-site seed sources) [198]
FIRE ADAPTATIONS AND PLANT RESPONSE TO FIRE
Desert-willow sprouts after top-kill by fire, and it may also establish from seed. Most riparian species are adapted to sprouting after damage from flooding; similarly, aboveground damage from fire can also trigger production of new shoots [204]. Desert-willow typically sprouts from the root crown after top-kill from low- to moderate-severity fire [12,18,218,219,244]. Postfire sprout mortality may be high during hot, dry periods and with lowering water tables [243]. Because the seeds are wind- and water-dispersed and have a transient, soil-stored seed bank, desert-willow may establish from seed after fire on moist sites (see Regeneration Processes).
Desert-willow occurs in early postfire environments [12,218,219,245]. Following a July wildfire in sugar sumac-coastal sage scrub oak-hairy yerba santa chaparral in San Diego County, California, more than 90% of desert-willows survived. These survivors sprouted by postfire month 2. They developed a multistemmed growth form and averaged 171 sprouts/plant by postfire month 10 (table 3) [219].
| Table 3—Postfire growth of desert-willow after the July 1975 San Ysidro chaparral wildfire in Anza-Borrego Desert State Park, California. Data are means. Adapted from Tratz (1978) [219]. | ||||
| Postfire month | Plants/ha | Sprouts/plant | Sprout length (cm) |
Aboveground mass/plant (g) |
| 2 (Sept.) | 30 | 16 | 5.5 | 2 |
| 4 (Nov.) | 10 | 21 | 27.1 | 40 |
| 7 (Feb.) | 30 | 48 | 47.6 | 132 |
| 10 (June) | 18 | 171 | 49.3 | 892 |
In 2003–2004 and again in 2012–2013, Bock and Bock (2014) measured the condition of riparian trees at the Appleton-Whittell Research Ranch, Arizona, in three canyons above the San Pedro River that had experienced wildfire in 2002, 2009, or both. Fire killed some desert-willows, but most were top-killed and sprouted. For desert-willows on sites that burned once, 10% survived as mature trees, 5% died, and 85% were partially to completely top-killed and sprouted. For desert-willows on sites that burned once or twice, 15% survived as mature trees, 20% died, and 65% were partially to completely top-killed and sprouted [12]. See the Research Project Summary of this study for more information on the response of desert-willow and associated trees (Arizona sycamore, Arizona walnut, Fremont cottonwood, and velvet ash) to these fires.
FUEL CHARACTERISTICS
Historically, fuels in low-elevation deserts of the Southwest were likely mostly sparse and discontinuous, although information on this is limited [60,109,243]. Keane et al. (2000) provide crown bulk density estimates for various structural stages of riparian forests and several other plant communities on the Gila National Forest. See the publication for further information [99].
Desert-willow communities typically have a moderate to deep litter layer, although litter decay rates may be fast. Desert-willow communities codominated by willow typically have deep litter, mostly due to the accumulation of willow leaves [155]. However, on the Jornada Experimental Range, New Mexico, sites dominated by desert-willow had loose litter that averaged 1 cm thick [95]. Over 6 months in Las Cruces, New Mexico, desert-willow leaf litter decayed faster on the soil surface than when buried 10 cm deep. Both surface and buried litter decayed faster in summer than in winter or spring. Schaefer et. al (1985) present a model for predicting rates of litter decay for desert-willow and associated shrub species [179].
Information on the flammability of desert-willow was lacking in the literature, but a few publications provide measures of live desert-willow fuels. Leaf area index is used in fuel models to help predict fire behavior and severity [212]. Studies in the Chiricahua Desert (location not provided) found desert-willow had lower mean specific leaf area (4.30 m²/kg) than surrounding desert shrub species [231], suggesting that it has less canopy fuel than some associated species. In southern New Mexico, surface area of desert-willow leaves averaged 3.1 cm², and specific leaf mass averaged 12.5 mg/cm² [102].
Climbing ladder fuels are present on some sites with desert-willow. For example, canyon grape often occurs in communities where desert-willow is dominant or important [7,140,207], and it can grow into upper canopies. At the Fort Bowie National Historic Site, Arizona, canyon grape formed continuous fuels from surface vegetation to the upper canopy of an Arizona walnut-netleaf hackberry-gum bully riparian woodland in which desert-willow was an associate [239].
Nonnative species can alter fuel distributions and loads. When riparian vegetation composition and structure change as a result of nonnative species in the overstory and/or understory, fuel characteristics may also change. In turn, this can alter fire intensity, severity, and/or rates of spread [243] (see Fire Regimes and Management Under a Changing Climate). Near Yuma, Arizona, fine fuels helped carry and spread the 2005 King Valley Fire, which burned a desert ironwood-blue paloverde-yellow paloverde-desert-willow community. The fine fuels were mostly Mediterranean grass (nonnative annual grasses) and desert Indianwheat (a native annual forb). These herbs had grown at high densities in response to the extremely wet winter of 2004–2005 [221].
FIRE REGIMES
In shrub and mixed-deciduous riparian communities of the Southwest and northern Mexico, wildfires are ignited by lightning in June and July [9]. Contemporary fires often spread into riparian areas from adjacent uplands [12,40], but fires can also start in riparian zones [243]. It is uncertain how frequent fires were within riparian areas of these deserts historically. Fires in small drainages likely had different frequencies and behaviors than fires in larger drainages where riparian zones were wider and vegetation more developed [12,160]. Fire intervals may be long and severity low in washes and riparian mixed-deciduous woodlands where periodic flooding scours the understory, limiting buildup of fine fuels and litter [243,244].
Information on historical fire regimes in riparian areas of the Southwest is limited due to lack of data [72,191,244], and the variability of historical fire regimes within and among desert riparian areas is not well known. The dynamic nature of riparian ecosystems makes their fire regimes hard to characterize [243,244]. Historically, sparse surface fuels and the variable patchiness of desert plant communities probably limited fire size, severity, and intensity, particularly in riparian zones [60,109,243]. Webb (2017, 2019) suggests that fires varied in severity and extent across southwestern riparian ecosystems [243,244]. Most fires were likely patchy and of low severity due to the patchiness of vegetation in desert landscapes; however, mixed- and high-severity fires likely occurred as well [111,243,244]. Fire might have been historically uncommon in some riparian areas [60,72,192,194,243,244]. In many cases, fires that started in the uplands might have slowed or stopped in riparian zones because temperatures were cooler and live fuel moistures higher [18,60,72,243]. Brooks et al. (2018) report that prior to the 1940s, ranchers who tried to burn in desert shrublands and Joshua tree woodlands were mostly unsuccessful due to fuel discontinuity [18].
In many areas, stand structure and function of mixed-deciduous riparian woodlands have been significantly altered since European-American exploration and settlement began [243]. Before that time, American Indians likely burned riparian zones, although the extent of such burning is unknown. Archeological studies along the Salt-Gila Aqueduct in southern Arizona revealed desert-willow charcoal deposits near prehistorical Hohokam settlements, and the researchers suggested these deposits were remnants of prehistorical agricultural burning [132,133]. Riparian areas experienced rapid ecological change [47,243,244] during European-American exploration (1540-1821) and settlement (1822-1912) periods [47]. In Arizona, for example, Spanish explorers imported livestock to the San Pedro Valley as early as the 1500s; European-Americans cut timber throughout the settlement period; miners diverted water from the San Pedro River to process ore in the 1800s; and fur trappers hunted American beavers until the animals were extirpated from the valley [243,244].
Changes in hydrology and vegetation have undoubtedly affected fire regimes of southwestern riparian communities and the surrounding uplands. However, the degree to which fire regimes are changing—or what the effects might be for riparian ecosystem structure, function, and resilience—are not well known [243,244]. The greatest ecological change in riparian areas since large-scale livestock ranching was introduced is probably a shift in fire regimes from mostly patchy surface fires to larger crown fires [5,243]. Some research suggests that fire frequency and severity are increasing in some southwestern riparian ecosystems [26,27,59,62,72,90,204,206,244], although the extent of these increases are largely unknown. Wildfire became a significant disturbance agent in southwestern riparian areas at the end of the 20th century. With the reduction in frequency and magnitude of floods, litter and debris accumulated in the forest understory. This accumulation, along with increased density of native and nonnative vegetation, resulted in greater fuel loads, fire sizes, and fire intensities than existed prior to riparian modification (review by [192]).
Factors that can increase fire severity include drought, altered flood disturbance, anthropogenic ignitions in high-use areas [18,26,59,62,206,243,250], flood and fire exclusion and associated high fuel loads, and more continuous fuels [18,59,62,72,77,243,250] resulting, in part, from spread of nonnative plant species [18,26,59,206,243]. Severe or intense fires can induce uniform changes in structure and composition across riparian zones by causing widespread mortality of native trees [8,243], increasing the likelihood of postfire establishment of nonnative plants [72,243,244]. Severe fires may be less common in areas with natural flood regimes [62,243,244].
Drought and/or human engineering can cause changes in hydrology (e.g., surface flows, groundwater levels, and channel and floodplain morphology) that favor nonnative plant species over native ones [27,59,72,90,169,244,257]. Nonnative annual grasses—particularly red brome, Mediterranean grass, and cheatgrass [17,18]—and nonnative trees Russian-olive [256] and tamarisk (saltcedar and five-stamen tamarisk [17,255], and potentially Athel tamarisk [234] and French tamarisk [29]) can increase fuel continuity and fire frequency in riparian communities of the Southwest [17,18] (see Fire Management Considerations). Fires may be larger, more frequent, and more severe in invaded communities. For example, fire-induced mortality of native riparian trees can increase dramatically when tamarisk occurs in the prefire plant community [244]. On 30 sites across the Southwest, high fire consumption and postfire mortality of Fremont cottonwoods and willows were positively associated with high prefire tamarisk cover. Fire was more severe where tamarisk was present than where it was not, resulting in more tissue damage to native trees. When prefire tamarisk cover was >50%, fire completely consumed the fine tissues of all native trees present [59].
Presettlement fire regimes of mixed-deciduous riparian communities of the Southwest likely varied among sites [243,244]. LANDFIRE (2008) models classify fire regimes of Biophysical Settings dominated by cottonwood-willow communities in the Southwest into two groups: Fire Regime Group I (<35-year return intervals, mixed severity) and Fire Regime Group V (>200-year fire return intervals, replacmente severity). Communities in the frequent-fire group are generally those along larger rivers, and those in the infrequent fire group are generally smaller desert riparian areas (i.e., stringers) [111].
See these FEIS publications for information on historical fire regimes in plant communities in which desert-willow is common to dominant: See table A3 for a list of additional plant communities in which desert-willow is common to dominant in wash zones, and search FEIS for fire regime information on those communities.FIRE MANAGEMENT CONSIDERATIONS
There is limited research on the impacts of fire on southwestern riparian communities [244]. Dams, waterway diversions, and climate change have greatly altered the hydrology of many streams and rivers. In the absence of natural flooding and with lowering water tables, fire is becoming the dominant disturbance in some riparian ecosystems of the Southwest. Groundwater depth and streamflow greatly influence vegetation type and extent in riparian ecosystems [26,77,90,243], and fire regimes seem to be changing in some lowland riparian communities with altered hydrology. Changing fire regimes are likely to have drastic, potentially irreversible effects on biodiversity and function of affected riparian communities. Alterations in plant community structure and plant and animal community composition may occur, including declines in wildlife species of concern [244]. Desert-willow may not survive severe, intense, or very frequent fire [12].
More severe, intense, and/or frequent fire can interact with altered hydrology to accelerate the replacement of native plants with nonnative plants [118,243]. In many riparian areas in the Southwest, there have been increases in tamarisk, Russian-olive, Mediterranean grass, and other nonnative plants that have different fuel characteristics, fire tolerances, and postfire fire responses than native species. Establishment and spread of tamarisk [170,255] and other nonnative species have changed fuel and fire regime characteristics in many riparian areas, and increased fire risk has been associated with high cover of tamarisk and Russian-olive [26,59,142,169,174,243,244]. Where groundwater levels have dropped, tamarisk has a competitive advantage over cottonwoods [118,243]. Its higher tolerance to drying soils [196] and frequent fire are driving changes in riparian stand structure and fire regimes, resulting in decreasing cover of formerly dominant, native trees such as desert-willow, cottonwoods, and willows [244,255]. Fire intervals in mixed-deciduous woodlands may be as frequent as 15 years in areas where tamarisk cover is high [59,110,163,243,244]. Where natural flow regimes are more intact and tamarisk is suppressed by native vegetation, desert-willow and other native trees are more likely to dominate postfire landscapes [244].
Fire management in southwestern riparian areas may include fuel reduction and prescribed fire treatments that promote or maintain native vegetation [244] and reduce or eradicate invasives [187,243]. Although simulated fuel reduction treatments showed potential to mitigate fire severity in some mixed-deciduous woodlands and forests under projected climate change in the Huachuca Mountains, projected basal area and mortality rates in riparian zones were unchanged by simulated fuel treatments, and a reduction in basal area and spatial extent of riparian species occurred with or without fire and regardless of treatments [151]. Prescribed fire may be most effective when used in combination with other restoration, such as restoring natural waterway flows [244]. However, little information was available on using prescribed fire in riparian plant communities of the Southwest [9]. The effects of fire, including prescribed burning, have been better studied in southwestern grasslands and savannas than in adjacent riparian woodlands and forests [12,60,72,191], so more information is needed on the use of prescribed fire and postfire restoration in riparian areas. Fire may be severe, and lethal to overstory deciduous trees, in riparian woodlands that have not experienced fire for many decades.
Bock and Bock (1990) do not recommend prescribed fire in late-successional riparian woodlands of the Southwest because "fire is difficult to manage and potentially very destructive" in those habitats [11]. Refined fuel and fire behavior models are needed for riparian ecosystems of the Southwest, including models for areas in which tamarisk has become dominant. Webb et al. (2019) review the information available for using prescribed fire in southwestern riparian areas, with and without invasive species [244].OTHER STATUS
Desert-willow shrublands are ranked Vulnerable (G3, S3) [144,146,159]. Other information on state- and province-level protection status of plants in the United States and
Canada is available at NatureServe.
IMPORTANCE TO WILDLIFE AND LIVESTOCK
A myriad of wildlife species use riparian habitats [48,49,81,124,134,237,241,244]. Riparian areas comprise a small part of total land area in the southwestern United States and northern Mexico, but they are extremely important to biotic communities [244]. Riparian areas are among the most diverse and productive ecosystems in desert bioregions, supporting floras and faunas that are biologically richer than adjacent uplands [21,32,35,244]. Riparian areas and washes provide water, cover, and breeding habitat for many species of wildlife, including obligate riparian species [70,134]. For wildlife that also use uplands, riparian areas provide refuge from the harsher, surrounding environments [134]. Riparian areas and washes also provide water, cover, and breeding habitat for livestock [24,70,134].
Wildlife that use riparian ecosystems for breeding, migration, and wintering habitat include large ungulate [48,244], small mammal, bird [2,48,81,168,205,241], reptile, amphibian, and arthropod species [22,25,48,168,244]. For example, On the Stockton Plateau of Texas, Arizona walnut-desert-willow communities provide habitat for white-tailed deer, ringtails, eastern fox squirrels, and Merriam's pocket mice [86]. Western white-throated woodrats along the Rio Grande use desert-willow wash habitats [253]. On the New Mexico State University's Agricultural Experiment Station, most (54%) of the litter and soil fauna collected under mature desert-willows were mites. Springtails were also common [95].
Riparian ecosystems provide habitat for many threatened or endangered species. Of 40 Federally listed Threatened or Endangered wildlife species in New Mexico—which includes several endemic species—at least 70% require aquatic and/or riparian habitat to feed, reproduce, and/or carry out their life cycles [222,244]. See these sources for checklists of birds that may use riparian communities with desert-willow for breeding and nesting habitat: [2,81,168,197,205]. Reynolds (1964) provides checklists of mammals and fish that may use riparian/aquatic habitats with desert-willow [168].
Communities with desert-willow provide important breeding, nesting, and migrant habitat for a wide variety of bird species [48,104,197,205]. For example, cactus wrens use desert-willow habitats for nesting and the fluffy desert-willow seeds for nesting material [135]. On the Barry M. Goldwater Air Force Range, southeastern Arizona, species richness of breeding and passing neotropical migrants is positively associated with high cover of mature (>2.5 m) desert-willow, paloverde, mesquite, and catclaw acacia. Migrant hummingbirds use desert-willow for cover and its nectar as a food source [83].
Many wildlife species browse desert-willow or consume its nectar or litter [22,218]. Large ungulates browse it, although their use may be limited. On the Elephant Mountain Wildlife Management Area in the Chihuahuan Desert, Texas, female bighorn sheep only browsed desert-willow in winter, although males browsed it year-round [15]. Desert-willow leaves and fruit provide minor forage for desert mule deer from spring through fall [25,103,106,186]. Various birds, including Gambel's quail [79,80], eat the seeds [232]. The sucrose in desert-willow nectar is a good energy source [22]. Hummingbirds and various insects, particularly bumble bees, consume the nectar [22]. Sonoran desert tortoises eat the fallen flowers [25]. Fly species feed from the extrafloral nectaries on the leaves [157]. In the Mojave Desert in southern Nevada, Cooper thrips live in and feed on decaying desert-willow litter [250].
Palatability and Nutritional Value
Desert-willow is not highly palatable to wildlife and livestock [45,100,232] because the foliage and bark contain glycosides and phenolic acids [138,252]. Substantial use typically occurs only when more palatable forage is not available [100,232], although wildlife may browse new sprouts after fire [219] or other top-killing disturbances.
See these sources for information on the nutritional value of desert-willow twigs: [33], leaves: [102] and seeds: [94,228].
Cover Value
Plant communities with desert-willow provide cover for many wildlife species [108,127]. They provide birds with hiding, resting, and nesting sites and foraging opportunities, some of which are absent or rare in adjacent plant communities [192]. Bird species using these communities include ferruginous pygmy-owls [173], northern bobwhites [82,88], Gambel’s quail [79,93], road runners [93], ladder-backed woodpeckers [93,95], hummingbirds [83], and many passerines [93] (e.g., gray vireos [249] and summer tanagers [205]).
High-quality fish habitat depends on suitable water temperatures for spawning and survival [48,78]. Desert-willow provides shade cover for fish, including the Federally Endangered [225] Rio Grande silvery minnow [41], and the Apache and Gila trout [48].
Burned areas create habitat for some wildlife guilds. They tend to increase hunting opportunities for predators that prefer hunting in open areas such as coyotes [36], common black hawks [178], and peregrine falcons [192]. Mixed-severity and severe fires create wildlife snags for cavity-nesting species [192].
VALUE FOR REHABILITATION OR RESTORATION
Desert-willow is used for erosion control [91,98,100,119,127], stream stabilization, and revegetation [89,127]. Because of the relatively fast growth rates of desert-willow and other riparian species, riparian communities generally recover faster than upland plant communities after restoration is implemented [134].
Desert-willow is propagated from seed, cuttings [58,119,127,220], and mound layering [57]. Cultivars are available [105,189,216,220]. See these sources for information on propagating desert-willow: [37,105,127].
OTHER USES
Riparian areas with desert-willow provide many ecosystem services including recreational opportunities, harboring pollinators, providing livestock forage [190,244], and enhancing water quality [244]. Riparian vegetation reduces nonpoint source pollution in waterways by filtering sediments, chemicals, and nutrients from overflow. The roots of streamside plants help prevent excessive erosion during floods [223] and promote groundwater infiltration [66], and vegetation surrounding active channels helps slow water velocity during floods [223]. Riparian trees and shrubs shade surface water, reducing water temperatures [223] and lost to evaporation [244].
Several products or potential products involve desert-willows. Desert-willow is planted as an ornamental [43,63,100,127,194,195,220] and used in shelterbelts [82]. The flowers are a bee food from which good honey is made [232]. Desert-willow can uptake gold from gold-enriched media, so it has potential as a gold mining plant [175]. Desert-willow tissues are being investigated for use as medicines. The foliage and bark have antibacterial and antifungal properties [138]. Fatty acids in the seeds may have anticarcinogenic and antidiabetic properties, as well as properties that may promote immune system and heart health [228].
Desert-willow wood provides timber, fenceposts, and firewood [91,98,119,148,161,220,232]. The wood is durable but not strong [91], and it has no commercial value [237]. See these sources for information on the properties of desert-willow wood: [152,153].
Native peoples have many traditional uses for desert-willow. They use the wood for house frames, granaries, and bows; the fibrous bark for weaving nets, shirts, and breechcloth [127]; and the branches for making baskets [220,232] and bows [188]. The flowers and seed capsules are eaten [127].
ADDITIONAL MANAGMENT CONSIDERATIONS
The value of southwestern riparian woodlands for both biotic communities and human livelihoods is well established [48,49,87,143,192,201,229], and maintaining and/or restoring riparian areas of the Southwest is a critical management issue [48,49,131,192]. Management goals for southwestern riparian areas may include floodplain restoration, safeguarding high-quality riparian areas, fuel reduction treatments, preventing spread of fire, and preventing establishment and spread of nonnative invasive plants [187,243].
Many riparian communities of the Southwest are highly imperiled due to anthropogenic disturbances [190]. Few naturally functioning riparian areas remain in the region, and those that do are threatened with climate change [134,244]. Within the last 100 to 150 years, humans have modified many riparian areas extensively by irrigating, pumping groundwater, building dams and diversions, clearing floodplains, cutting wood, OHV use [69], grazing livestock [23], and spreading nonnative invasive plants. These modifications have resulted in significant changes in the physical environment and biota [64,134,136,202,244]. For example, livestock generally use riparian areas more than adjacent, less productive upland communities, compacting soils and reducing recruitment of palatable deciduous seedlings and saplings into the overstory [44,134]. Use of the relatively unpalatable desert-willow by livestock generally indicates overbrowsing or overstocking [227,240].
Altered hydrological regimes are often associated with establishment and spread of nonnative plants. Altered hydrology and sediment deposition resulting from river impoundment may reduce or eliminate germination and recruitment of native deciduous trees while promoting germination and recruitment of tamarisk [244]. Consequently, tamarisk species have established in many riparian areas of the Southwest [4,134,170,203,204,255], including wash and woodland communities in which desert-willow was formerly dominant or important [20,123,125] (see Fire Management Considerations). Maintaining an ecologically healthy riparian community can help stop establishment and spread of tamarisk and other invasive plant species [3]. Dense shade in closed-canopy communities with desert-willow may inhibit establishment of tamarisk [52].
Flow regimes that fall within the natural range of variability for undammed waterways can have multiple benefits, including:
MANAGEMENT UNDER A CHANGING CLIMATE
Southwestern riparian ecosystems are vulnerable to decline under hotter and drier conditions [31,201,243,244]. Climate change and variation—and associated impacts on hydrology, channel morphology, and riparian vegetation—may alter fire regimes by increasing the likelihood of more frequent or severe wildfires [12,60,193,244]. For example, prolonged drought increases fire risk by decreasing water availability and reducing fuel moisture [160,243,244].
Changes in water flow are expected with climate change, including lower water tables and increased frequency and severity of extreme weather that will likely result in more frequent and severe floods as well as more intense droughts [61,72,181,191,192,243]. Warming in montane areas may increase the number of severe floods downstream by increasing the rate of snowmelt into downstream waterways such as the Middle Rio Grande [243]. Monsoons may occur later in the year and with greater severity [192]. Droughts are likely to be increasingly severe and prolonged, resulting in further reductions in discharge volume [192].
Distribution and composition of upland communities may change with climate warming [70,243,244]. In turn, this may affect composition of riparian vegetation in ways that increase fire risk (e.g., facilitating spread of nonnative species [191]). Climate change could alter competitive relationships between desert-willow and other native species, and also between desert-willow and nonnative species such as tamarisk and Russian-olive [142]. For example, if flood timing does not coincide with spring dispersal of desert-willow, cottonwood, and willow seeds, reproductive success of these native trees is likely to decrease while that of nonnative trees is likely to increase [71,191].
| Table A1—Common and scientific names of plants mentioned in this review. Links go to FEIS Species Reviews. | |
| Common name | Scientific name |
| Cacti | |
| cactus | Cactaceae |
| Forbs | |
| desert Indianwheat | Plantago ovata |
| Graminoids | |
| Arizona muhly | Muhlenbergia arizonica |
| blue grama | Bouteloua gracilis |
| cheatgrassa | Bromus tectorum |
| Mediterranean grassa | Schismus spp. |
| Hall's panicgrass | Panicum hallii |
| red bromea | Bromus rubens |
| sideoats grama | Bouteloua curtipendula |
| tobosagrass | Pleuraphis mutica |
| Lianas | |
| canyon grape | Vitis arizonica |
| eastern poison-ivy | Toxicodendron radicans |
| Shrubs | |
| Apache plume | Fallagia paradoxa |
| baccharis | Baccharis spp. |
| blackbrush | Coleogyne ramosissima |
| burrobush | Ambrosia dumosa |
| catclaw acacia | Senegalia greggii |
| coastal sage scrub oak | Quercus dumosa |
| creosote bush | Larrea tridentata |
| desertbroom | Baccharis sarothroides |
| desert ironwood | Olneya tesota |
| eastern Mojave buckwheat | Eriogonum fasciculatum |
| gum bully | Sideroxylon lanuginosa |
| hairy yerba santa | Eridictyon trichocalyx |
| honey mesquite | Prosopis glandulosa |
| netleaf hackberry | Celtis laevigata var. reticulata |
| mesquite | Prosopis spp. |
| paloverde | Parkinsonia spp. |
| screwbean mesquite | Prosopis pubescens |
| singlewhorl burrobrush | Hymenoclea monogyra |
| splitleaf brickellbush | Brickellia laciniata |
| smoketree | Psorothamnus spinosus |
| spiny hackberry | Celtis ehrenbergiana |
| sugar sumac | Rhus ovata |
| Texas paloverde | Parkinsonia texana |
| tree tobacco | Nicotiana glauca |
| velvet mesquite | Prosopis velutina |
| yellow paloverde | Parkinsonia microphylla |
| yucca | Yucca spp. |
| Trees | |
| alligator juniper | Juniperus deppeana |
| American sycamore | Platanus occidentalis |
| Arizona sycamore | Platanus wrightii |
| Arizona walnut | Juglans major |
| arroyo willow | Salix lasiolepis |
| Athel tamarisk | Tamarix aphylla |
| black willow | Salix nigra |
| blue paloverde | Parkinsonia florida |
| canyon live oak | Quercus chrysolepis |
| cottonwood | Populus spp. |
| desert-willow | Chilopsis linearis |
| five-stamen tamariska | Tamarix chinensis |
| Fremont cottonwood | Populus fremontii |
| French tamariska | Tamarix gallica |
| Goodding’s willow | Salix gooddingii |
| green ash | Fraxinus pennsylvanica |
| Joshua tree | Yucca brevifolia |
| juniper | Juniperus spp. |
| little walnut | Juglans microcarpa |
| Lombardy poplara | Populus nigra |
| narrowleaf willow | Salix exigua |
| oak | Quercus spp. |
| Parry pinyon | Pinus quadrifolia |
| Russian-olivea | Elaeagnus angustifolia |
| saltcedara | Tamarix ramosissima |
| singleleaf pinyon | Pinus monophylla |
| tamariska | Tamarix spp. |
| Texas madrone | Arbutus xalapensis |
| velvet ash | Fraxinus velutina |
| wavyleaf oak | Quercus × pauciloba |
| western soapberry | Sapindus saponaria var. drummondii |
| white alder | Alnus rhombifolia |
| willow | Salix spp. |
| aNonnative species. | |
| Table A2—Common and scientific names of wildlife species mentioned in this review. Links go to FEIS Species Reviews. | |
| Common name | Scientific name |
| Arthropods | |
| bees | Apoidea |
| bumble bee | Bombus spp. |
| carpenter bees | Xylocopinae |
| Cooper thrip | Hoplothrips cooperi |
| flies | Diptera |
| hawkmoth | Hyles spp. |
| honeybee | Apis mellifera |
| lady-beetles | Coccinellidae |
| mites | Trombidiformes |
| Sonoran bumble bee | Bombus sonorous |
| springtails | Collembola |
| Fish | |
| Apache trout | Oncorhynchus apache |
| Gila trout | Oncorhynchus gilae |
| Rio Grande silvery minnow | Hybognathus amarus |
| Reptiles | |
| Sonoran desert tortoise | Gopherus morafkai |
| Birds | |
| cactus wren | Campylorhynchus brunneicapillus |
| common black hawk | Buteogallus anthracinus |
| ferruginous pygmy-owl | Glaucidium brasilianum |
| Gambel’s quail | Callipepla gambelii |
| gray vireo | Vireo vicinior |
| ladder-backed woodpecker | Dryobates scalaris |
| road runner | Geococcyx spp. |
| hummingbirds | Trochilidae |
| northern bobwhite | Colinus virginianus |
| passerines | Passeriformes |
| summer tanager | Piranga rubra |
| Mammals | |
| American beaver | Castor canadensis |
| bighorn sheep | Ovis canadensis |
| desert mule deer | Odocoileus hemionus eremicus |
| eastern fox squirrel | Sciurus niger |
| Merriam's pocket mouse | Perognathus merriami |
| ringtail | Bassariscus astutus |
| western white-throated woodrat | Neotoma albigula |
| white-tailed deer | Odocoileus virginianus |
| Table A3—Representative plant community classifications in which desert-willow occurs. | |
| FRES Ecosystems | |
| FRES30 Desert shrub | |
| FRES32 Texas savanna | |
| FRES33 Southwestern shrubsteppe | |
| FRES 35 Pinyon-juniper | |
| FRES38 Plains grasslands | |
| FRES40 Desert grasslands [74] | |
| Kuchler Plant Associations | |
| K023 Juniper-pinyon woodland | |
| K027 Mesquite bosque | |
| K031 Oak-juniper woodlands | |
| K041 Creosotebush | |
| K042 Creosotebush -bursage | |
| K043 Paloverde-cactus shrub | |
| K044 Creosotebush-tarbush | |
| K054 Grama-tobosa prairie | |
| K058 Grama-tobosa shrubsteppe | |
| K059 Trans-Pecos shrub savanna | |
| K061 Mesquite-acacia savanna [107] | |
| SAF Cover Types | |
| 222 Black cottonwood-willow | |
| 235 Cottonwood-willow | |
| 239 Pinyon-juniper | |
| 242 Mesquite [65] | |
| SRM (Rangeland) Cover Types | |
| 203 Riparian woodland | |
| 211 Creosote bush scrub | |
| 212 Blackbush | |
| 412 Juniper-pinyon | |
| 422 Riparian | |
| 502 Grama-galleta | |
| 504 Juniper-pinyon pine | |
| 506 Creosotebush-bursage | |
| 507 Paloverde-cactus | |
| 508 Creosotebush-tarbush | |
| 707 Blue grama-sideoats grama-black grama | |
| 729 Mesquite | |
| 735 Sideoats grama-sumac-juniper [185] | |
1. Allred, Kelly W.; Jercinovic, Eugene M.; DeWitt, Ivey. 2020. Flora Neomexicana III: An illustrated identification Manual; Part 2: Dicotyledonous plants. 2nd ed. Independently published. 795 p. [94735]
2. Anderson, Bertin W.; Ohmart, Robert D. 1977. Vegetation structure and bird use in the Lower Colorado River Valley. In: Johnson, R. Roy; Jones, Dale A., tech. coords. Importance, preservation and management of riparian habitat: A symposium; 1997 July 9; Tucson, AZ. Gen. Tech. Rep. RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 23-34. [52882]
3. Anderson, Bertin. 1998. The case for salt cedar. Restoration and Management Notes. 16(2): 130-134. [44007]
4. Bagstad, K. J.; Lite, S. J.; Stromberg, J. C. 2006. Vegetation, soils, and hydrogeomorphology of riparian patch types of a dryland river. Western North American Naturalist. 66(1): 23-44. [62270]
5. Bahre, Conrad J. 1995. Human disturbance and vegetation in Arizona's Chiricahua Mountains in 1902. Desert Plants. 11(4): 41-45. [26028]
6. Baldwin, Bruce G.; Goldman, Douglas H.; Keil, David J.; Patterson, Robert; Rosatti, Thomas J.; Wilken, Dieter H., eds. 2012. The Jepson manual. Vascular plants of California, second edition. Berkeley, CA: University of California Press. 1568 p. [86254]
7. Bassett, Dick; Larson, Milo; Moir, Will. 1987. Forest and woodland habitat types (plant associations) of Arizona south of the Mogollon Rim and southwestern New Mexico. 2nd ed. Albuquerque, NM: U.S. Department of Agriculture, Forest Service, Southwestern Region. Variously paginated. [20308]
8. Bendix, Jacob; Cowell, C. Mark. 2010. Impacts of wildfire on the composition and structure of riparian forests in southern California. Ecosystems. 13(1): 99-107. [81845]
9. Bennett, Peter S.; Kunzmann, Michael R. 1992. The applicability of generalized fire prescriptions to burning of Madrean evergreen forest and woodland. Journal of the Arizona-Nevada Academy of Science. 24-25: 79-84. [18324]
10. Bertelsen, David C. 2018. Thirty-Seven years on a mountain trail: Vascular flora and flowering phenology of the Finger Rock Canyon Watershed, Santa Catalina Mountains, Arizona. Desert Plants. 34(1-2): 1-247. [95896]
11. Bock, Carl E.; Bock, Jane H. 1990. Effects of fire on wildlife in southwestern lowland habitats. In: Krammes, J. S., technical coordinator. Effects of fire management of southwestern natural resources: Proceedings of the symposium; 1988 November 15-17; Tucson, AZ. Gen. Tech. Rep. RM-191. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 50-64. [11273]
12. Bock, Carl E.; Bock, Jane H. 2014. Effects of wildfire on riparian trees in southeastern Arizona. The Southwestern Naturalist. 59(4): 568-574. [89514]
13. Boles, Patrick H.; Dick-Peddie, William A. 1983. Woody riparian vegetation patterns on a segment of the Mimbres River in southwestern New Mexico. The Southwestern Naturalist. 28(1): 81-87. [65317]
14. Bonner, Franklin. 2008. Seed biology. In: Bonner, Franklin T.; Karrfalt, Robert P., eds. Woody plant seed manual. Agric. Handbook No. 727. Washington, DC: U.S. Department of Agriculture, Forest Service: 3-38. [45117]
15. Brewer, Clay E.; Harveson, Louis A. 2007. Diets of bighorn sheep in the Chihuahuan Desert, Texas. The Southwestern Naturalist. 51(1): 97-103. [95949]
16. Briggs, Mark K. 1996. Water availability. In: Briggs, Mark K. Riparian ecosystem recovery in arid lands: Strategies and references. Tucson, AZ: The University of Arizona Press: 61-77. [44786]
17. Brooks, Matthew L. 2008. Plant invasions and fire regimes. In: Zouhar, Kristin; Smith, Jane Kapler; Sutherland, Steve; Brooks, Matthew L., eds. Wildland fire in ecosystems: Fire and nonnative invasive plants. Gen. Tech. Rep. RMRS-GTR-42, Vol. 6. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 33-45. [70467]
18. Brooks, Matthew L.; Minnich, Richard A.; Matchett, John R. 2018. Southeastern deserts bioregion. In: van Wagtendonk, Jan W.; Sugihara, Neil G.; Stephens, Scott L.; Thode, Andrea E.; Shaffer, Kevin E.; Fites-Kaufman, Jo Ann, eds. Fire in California's ecosystems. 2nd ed. Oakland, CA: University of California Press: 353-378. [93914]
19. Brown, David E., ed. 1982. Biotic communities of the American Southwest--United States and Mexico. Desert Plants: Special Issue. Tucson, AZ: University of Arizona Press. 4(1-4): 1-342. [62041]
20. Brown, David E.; Lowe, Charles H. 1974. A digitized computer-compatible classification for natural and potential vegetation in the Southwest with particular reference to Arizona. Journal of the Arizona Academy of Science. 9 (Suppl. 2): 3-11. [20374]
21. Brown, David E.; Lowe, Charles H.; Hausler, Janet F. 1977. Southwestern riparian communities: Their biotic importance and management in Arizona. In: Johnson, R. Roy; Jones, Dale A., tech. coords. Importance, preservation and management of riparian habitat: A symposium: Proceedings; 1977 July 9; Tucson, AZ. Gen. Tech. Rep. RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 201-211. [5348]
22. Brown, James H.; Kodric-Brown, Astrid; Whitham, Thomas G.; Bond, Hedley W. 1981. Competition between hummingbirds and insects for the nectar of two species of shrubs. The Southwestern Naturalist. 26(2): 133-145. [12236]
23. Brunelle, A.; Minckley, T. A.; Delgadillo, J.; Blissett, S. 2014. A long-term perspective on woody plant encroachment in the desert southwest, New Mexico, USA. Journal of Vegetation Science. 25(3): 829-838. [96267]
24. Buegge, J. Jeremy. 2001. Flora of the Santa Teresa Mountains in Graham County, Arizona. Journal of the Arizona-Nevada Academy of Science. 33(2): 132-149. [45078]
25. Bury, R. Bruce; Germano, David J.; Van Devender, Thomas R.; Martin, Brent E. 2002. The desert tortoise in Mexico. In: Van Devender, Thomas R., ed. The Sonoran desert tortoise: Natural history, biology, and conservation. Arizona-Sonora Desert Museum Studies in Natural History. Tucson, AZ: The University of Arizona Press; The Arizona-Sonora Desert Museum: 86-108. [69904]
26. Busch, David E. 1995. Effects of fire on southwestern riparian plant community structure. The Southwestern Naturalist. 40(3): 259-267. [26498]
27. Busch, David E.; Smith, Stanley D. 1993. Effects of fire on water and salinity relations of riparian woody taxa. Oecologia. 94(2): 186-194. [88320]
28. Calflora. 2022. The Calflora database: Information on California plants for education and conservation, [Online]. Berkeley, CA: Calflora (Producer). Available: http://www.calflora.org/. [94823]
29. California Invasive Pest Council. 2022. Cal-IPC: Plants A to Z. In: Cal-IPC, [Online]. Berkeley, CA: California Invasive Pest Council (Producer). Available: https://www.cal-ipc.org/plants/profiles/ [2020, November 22]. [94821]
30. Campbell, C. J.; Green, Win. 1968. Perpetual succession of stream-channel vegetation in a semiarid region. Journal of the Arizona Academy of Science. 5(2): 86-90. [53565]
31. Capon, Samantha J.; Chambers, Lynda E.; Mac Nally, Ralph; Naiman, Robert J.; Davies, Peter; Marshall, Nadine; Pittock, Jamie; Reid, Michael; Capon, Timothy; Douglas, Michael; Catford, Jane; Baldwin, Darren S.; Stewardson, Michael; Roberts, Jane; Parsons, Meg; Williams, Stephen E. 2013. Riparian ecosystems in the 21st century: Hotspots for climate change adaptation? Ecosystems. 16(3): 359-381. [96207]
32. Carothers, Steven W.; Johnson, R. Roy; Aitchison, Stewart W. 1974. Population structure and social organization of southwestern riparian birds. American Zoologist. 14(1): 97-108. [24192]
33. Carr, M. E.; Phillips, B. S.; Bagby, M. O. 1985. Xerophytic species evaluated for renewable energy resources. Economic Botany. 39(4): 505-513. [71932]
34. Carter, Jack L. 1997. Trees and shrubs of New Mexico. Boulder, CO: Johnson Books. 534 p. [72647]
35. Chaney, Ed; Elmore, Wayne; Platts, William S. 1990. Livestock grazing on western riparian areas. Eagle, ID: Northwest Resource Information Center, Inc. Produced for the U.S. Environmental Protection Agency. 45 p. [17790]
36. Chapman, Joseph A.; Feldhamer, George A., eds. 1982. Wild mammals of North America. Baltimore, MD: The Johns Hopkins University Press. 1147 p. [21085]
37. Chase, Valerie C.; Strain, B. R. 1966. Propagation of some woody desert perennials by stem cuttings. Madrono. 18(8): 240-243. [49002]
38. Clover, Elzada U.; Jotter, Lois. 1944. Floristic studies in the Canyon of the Colorado and tributaries. The American Midland Naturalist. 32(3): 591-642. [62472]
39. Cockman, Joneen S.; Pieper, Rex; Clason, Dennis. 1996. Arroyo-riparian shrub diversity along a transition zone between the Sacramento Mountains and Tularosa Basin, New Mexico. In: Barrow, Jerry R.; McArthur, E. Durant; Sosebee, Ronald E.; Tausch, Robin J., compilers. Proceedings: Shrubland ecosystem dynamics in a changing environment; 1995 May 23-25; Las Cruces, NM. Gen. Tech. Rep. INT-GTR-338. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 230-236. [27053]
40. Coffman, Gretchen C.; Ambrose, Richard F.; Rundel, Philip W. 2010. Wildfire promotes dominance of invasive giant reed (Arundo donax) in riparian ecosystems. Biological Invasions. 12(8): 2723-2734. [82396]
41. Coleman, Ross; Hutson, Alison M.; Toya, Louie A.; Tave, Douglas. 2011. Using native plants to provide natural ecosystem functions in a conservation fish hatchery. Native Plants Journal. 12(3): 216-225. [95950]
42. Cornell, Jack E.; Gutierrez, Melida; Wait, D. Alexander; Rubio-Arias, Hector O. 2008. Ecological characterization of a riparian corridor along the Río Conchos, Chihuahua, Mexico. The Southwestern Naturalist. 53(1): 96-100. [95953]
43. Cronquist, Arthur; Holmgren, Arthur H.; Holmgren, Noel H.; Reveal, James L.; Holmgren, Patricia K. 1984. Intermountain flora: Vascular plants of the Intermountain West, U.S.A. Vol. 4: Subclass Asteridae, (except Asteraceae). New York: The New York Botanical Garden. 573 p. [718]
44. Davis, Gary A. 1977. Management alternatives for the riparian habitat in the Southwest. In: Johnson, Roy; Jones, Dale A., technical coordinators. Importance, preservation and management of riparian habitat: A symposium; 1977 July 9; Tucson, AZ. General Technical Report RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 59-67. Available from: NTIS, Springfield, VA 22151; PB-274 582. [5336]
45. Dayton, William A. 1931. Important western browse plants. Misc. Publ. No. 101. Washington, DC: U.S. Department of Agriculture. 214 p. [768]
46. de Soyza, Amrita G.; Killingbeck, Keith T.; Whitford, Walter G. 2004. Plant water relations and phytosynthesis during and after drought in a Chihuahuan desert arroyo. Journal of Arid Environments. 59(1): 27-39. [48568]
47. DeBano, Leonard F.; Ffolliott, Peter F. 2002. Riparian history of the Southwest. In: Hydrology and water resources in Arizona and the Southwest: Proceedings, Arizona-Nevada Academy of Science--Hydrology section; 2002 April 6; Glendale, AZ. Volume 32. Mesa, AZ: Arizona-Nevada Academy of Science: 33-38. [45005]
48. DeBano, Leonard F.; Rinne, John N.; Baker, Malchus B., Jr. 2003. Management of natural resources in riparian corridors. Journal of the Arizona-Nevada Academy of Science. 35(1): 58-70. [96273]
49. DeBano, Leonard F.; Schmidt, Larry J. 1989. Improving southwestern riparian areas through watershed management. Gen. Tech. Rep. RM-182. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 33 p. [11016]
50. DeBolt, Ann; Spurrier, Carol S. 2004. Seeds of success and the Millennium Seed Bank project. In: Hild, Ann L.; Shaw, Nancy L.; Meyer, Susan E.; Booth, D. Terrance; McArthur, E. Durant, compilers. Seed and soil dynamics in shrubland ecosystems: Proceedings; 2002 August 12-16; Laramie, WY. Proceedings RMRS-P-31. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 100-108. [49104]
51. DePree, Elaine; Ludwig, John A. 1978. Vegetative and reproductive growth patterns in desert willow (Chilopsis linearis (Cav.) Sweet). The Southwestern Naturalist. 23(2): 239-246. [12237]
52. Dewine, J. M.; Cooper, D. J. 2008. Canopy shade and the successional replacement of tamarisk by native box elder. Journal of Applied Ecology. 45(2): 505-514. [70231]
53. Dick-Peddie, William A. 1993. New Mexico vegetation: Past, present, and future. Albuquerque, NM: University of New Mexico Press. 244 p. [21097]
54. Dick-Peddie, William A.; Hubbard, John P. 1977. Classification of riparian vegetation. In: Johnson, R. Roy; Jones, Dale A., tech. coords. Importance, preservation and management of riparian habitat: A symposium: Proceedings; 1977 July 9; Tucson, AZ. Gen. Tech. Rep. RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 85-90. [5338]
55. Diggs, George M., Jr.; Lipscomb, Barney L.; O' Kennon, Robert J. 1999. Illustrated flora of north-central Texas. Sida Botanical Miscellany, No. 16. Fort Worth, TX: Botanical Research Institute of Texas. 1626 p. [35698]
56. Donart, Gary B.; Sylvester, Donell; Hickey, Wayne. 1978. A vegetation classification system for New Mexico, U.S.A. In: Hyder, Donald N., ed. Proceedings, 1st international rangeland congress; 1978 August 14-18; Denver, CO. Denver, CO: Society for Range Management: 488-490. [4094]
57. Dreesen, David R.; Harrington, John T. 1997. Propagation of native plants for restoration projects in the southwestern U.S.--preliminary investigations. In: Landis, Thomas D.; Thompson, Jan R., tech. coords. National proceedings: Forest and conservation nursery associations--1997; Regeneration, reforestation, restoration: The seedling is the key; 1997 August 11-14; Bemidji, MN; 1997 August 19-21; Boise, ID. Gen. Tech. Rep. PNW-GTR-419. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 77-88. [29122]
58. Dreesen, David; Harrington, John; Subirge, Tom; Stewart, Pete; Fenchel, Greg. 2002. Riparian restoration in the Southwest: Species selection, propagation, planting methods, and case studies. In: Dumroese, R. Kasten; Riley, Lee E.; Landis, Thomas D., tech. coords. National proceedings: Forest and conservation nursery associations 1999, 2000, and 2001; [Multiple dates]; [Multiple locations]. Proceedings RMRS-P24. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 253-272. [47622]
59. Drus, Gail Michelle. 2013. Tamarisk (Tamarix spp.) and desert riparian ecosystem change. Santa Barbara, CA: University of California, Santa Barbara. 173 p. Dissertation. [88329]
60. Dwire, Kathleen A.; Kauffman, J. Boone. 2003. Fire and riparian ecosystems in landscapes of the western USA. In: Young, Michael K.; Gresswell, Robert E.; Luce, Charles H., eds. Selected papers from an international symposium on effects of wildland fire on aquatic ecosystems in the western USA; 2002 April 22-24; Boise, ID. In: Forest Ecology and Management. 178(1-2): 61-74. [44923]
61. Easterling, David R.; Meehl, Gerald A.; Parmesan, Camille; Changnon, Stanley A.; Karl, Thomas R.; Mearns, Linda O. 2000. Climate extremes: Observations, modeling, and impacts. Science. 289(5487): 2068-2047. [96228]
62. Ellis, Lisa M. 2001. Short-term response of woody plants to fire in a Rio Grande riparian forest, central New Mexico, USA. Biological Conservation. 97(2): 159-170. [38945]
63. Erdman, Kimball S. 1970. Distribution of the native trees of Utah. Brigham Young University Science Bulletin: Biological Series. 11(3): 1-34. [35781]
64. Espinoza, Jose Javier Ochoa; Ayala, Cesar Cantu; Castillon, Eduardo Estrada; Saldivar, Fernando Gonzalez; Sauceda, Jose Uvalle; Jurado, Enrique; ChapaVargas, Leonardo; Jaramillo, Edmar Melendez; Hernandez, Edgardo Ortiz. 2017. Livestock effect on floristic composition and vegetation structure of two desert scrublands In Northwest Coahuila, Mexico. The Southwestern Naturalist. 62(2): 138-145. [95955]
65. Eyre, F. H., ed. 1980. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters. 148 p. [905]
66. Ffolliott, P. F.; Baker, M. B., Jr.; DeBano, L. F.; Neary, D. G. 2004. Introduction. In: Baker, Jr., Malchus B.; Ffolliott, Peter F.; DeBano, Leonard F.; Neary, Daniel G., eds. Riparian areas of the southwestern United States: Hydrology, ecology, and management. Washington, DC: Lewis Publishers: 1-8. [96192]
67. Ffolliott, Peter F.; Thorud, David B. 1974. Vegetation for increased water yield in Arizona. Tech. Bull. 215. Tucson, AZ: University of Arizona, Agricultural Experiment Station. 38 p. [4448]
68. Fosberg, F. Raymond. 1936. Varieties of the desert willow, Chilopsis linearis. Madrono. 3(8): 362-366. [95956]
69. Frank, T. D.; Tweddale, S. A. 2006. The effect of spatial resolution on measurement of vegetation cover in three Mojave Desert shrub communities. Journal of Arid Environments. 67(Supplement): 88-99. [95958]
70. Friggens, M., Loehman, R., Holsinger, L., Finch, D. 2014. Vulnerability of riparian obligate species to the interactive effect of fire, climate and hydrological change: Final report. Interagency Agreement #13-IA-11221632-006. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. Available: https://www.fs.usda.gov/rm/pubs_journals/2014/rmrs_2014_friggins_m002.pdf. 213 p. [96213]
71. Friggens, Megan M.; Finch, Deborah M.; Bagne, Karen E.; Coe, Sharon J.; Hawksworth, David L. 2013. Vulnerability of species to climate change in the Southwest: Terrestrial species of the Middle Rio Grande. Gen. Tech. Rep. RMRS-GTR-306. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 191 p. [96212]
72. Friggens, Megan M.; Matthews, Stephen N. 2012. Risk assessment for two bird species in northern Wisconsin. In: Vose, James M.; Peterson, David L.; Patel-Weynand, Toral, eds. Effects of climatic variability and change on forest ecosystems: A comprehensive science synthesis for the U.S. forest sector. PNW-GTR-870. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: 256-258. [87089]
73. Gardner, J. L. 1951. Vegetation of the creosotebush area of the Rio Grande Valley in New Mexico. Ecological Monographs. 21(4): 379-403. [4243]
74. Garrison, George A.; Bjugstad, Ardell J.; Duncan, Don A.; Lewis, Mont E.; Smith, Dixie R. 1977. Vegetation and environmental features of forest and range ecosystems. Agric. Handb. 475. Washington, DC: U.S. Department of Agriculture, Forest Service. 68 p. [998]
75. Genoways, Hugh H.; Baker, Robert J.; Cornely, John E. 1979. Mammals of the Guadalupe Mountains National Park, Texas. In: Genoways, Hugh H.; Baker, Robert J., eds. Biological investigations in the Guadalupe Mountains National Park: Proceedings of a symposium; 1975 April 4-5; Lubbock, TX. Proceedings and Transactions Series No. 4. Washington, DC: U.S. Department of the Interior, National Park Service: 271-332. [16022]
76. Gibbens, Robert P. 1994. SRM 505: Grama-tobosa shrub. In: Shiflet, Thomas N., ed. Rangeland cover types of the United States. Denver, CO: Society for Range Management: 65 p. [67045]
77. Glenn, Edward P.; Nagler, Pamela L. 2005. Comparative ecophysiology of Tamarix ramosissima and native trees in western U.S. riparian zones. Journal of Arid Environments. 61(3): 419-446. [51922]
78. Gresswell, Robert E. 1999. Fire and aquatic ecosystems in forested biomes of North America. Transactions of the American Fisheries Society. 128(2): 193-221. [31018]
79. Gullion, Gordon W. 1960. The ecology of Gambel's quail in Nevada and the arid Southwest. Ecology. 41(3): 518-536. [49039]
80. Gullion, Gordon W. 1964. Wildlife uses of Nevada plants. Contributions toward a flora of Nevada: No. 49. CR-24-64. Beltsville, MD: U.S. Department of Agriculture, Agricultural Research Service, Crops Research Division; Washington, DC: U.S. National Arboretum, Herbarium. 170 p. [6729]
81. Hall, Linnea S.; Morrison, Michael L.; Christoferson, Laurel L.; Martin, John; Bock, Carl E.; Strong, Thomas R. 2002. Bird populations in riparian areas of southeastern Arizona in 1985-1986 and 1994-1995. Western North American Naturalist. 62(3): 370-376. [43921]
82. Hanson, William R. 1957. Plants for improving bobwhite habitat in northwestern Oklahoma. Arts and Sciences Studies: Biological Series Publication No. 7. Stillwater, OK: Oklahoma State University. 88 p. [62426]
83. Hardy, Paul C.; Griffin, David J.; Kuenzi, Amy J.; Morrison, Michael L. 2004. Occurrence and habitat use of passage neotropical migrants in the Sonoran Desert. Western North American Naturalist. 64(1): 59-71. [62255]
84. Harris, Lisa K.; Ruther, Sherry. 2000. Ecological characteristics of riparian washes in southeastern Arizona, USA. Natural Areas Journal. 20(3): 221-226. [35751]
85. Henrickson, James. 1985. A taxonomic revision of Chilopsis (Bignoniaceae). Aliso. 11(2): 179-197. [12058]
86. Hermann, Jack A. 1950. The mammals of the Stockton Plateau of northeastern Terrell County, Texas. The Texas Journal of Science. 2(3): 368-393. [61365]
87. Howe, W. H.; Knopf, F. L. 1991. On the imminent decline of Rio Grande cottonwoods in central New Mexico. The Southwestern Naturalist. 36(2): 218-224. [18278]
88. Jackson, A. S. 1969. A handbook for bobwhite quail management in the West Texas Rolling Plains. Bulletin No. 48. [Austin, TX]: Texas Parks and Wildlife Department. 77 p. [62367]
89. James, Richard D. 1998. Use of native species in revegetation of disturbed sites (Arizona). In: Tellman, Barbara; Finch, Deborah M.; Edminster, Carl; Hamre, Robert, eds. The future of arid grasslands: Identifying issues, seeking solutions: Proceedings; 1996 October 9-13; Tucson, AZ. Proceedings RMRS-P-3. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 297-303. [29296]
90. Jemison, Roy. 2004. Relationships between hydrology, exotic plants, and fuel loads in the Middle Rio Grande. In: Hydrology and water resources in Arizona and the Southwest: Proceedings, Arizona-Nevada Academy of Science--Hydrology section; 2003 April 12; Flagstaff, AZ. Volume 33. Glendale, AZ: Arizona Nevada Academy of Science: 85-92. [52856]
91. Johnson, Carl M. 1970. Common native trees of Utah. Special Report 22. Logan, UT: Utah State University, College of Natural Resources, Agricultural Experiment Station. 109 p. [9785]
92. Johnson, Hyrum B. 1976. Vegetation and plant communities of southern California deserts--a functional view. In: Latting, June, ed. Symposium proceedings: Plant communities of southern California; 1974 May 4; Fullerton, CA. Special Publication No. 2. Berkeley, CA: California Native Plant Society: 125-164. [1278]
93. Johnson, R. Roy; Haight, Lois T.; Riffey, Meribeth M.; Simpson, James M. 1980. Brushland/steppe bird populations. In: DeGraaf, Richard M., technical coordinator. Workshop proceedings: Management of western forests and grasslands for nongame birds; 1980 February 11-14; Salt Lake City, UT. Gen. Tech. Rep. INT-86. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 98-112. [17900]
94. Jones, Quintin; Barclay, A. S. 1972. Industrial raw materials from shrubs. In: McKell, Cyrus M.; Blaisdell, James P.; Goodin, Joe R., tech. eds. Wildland shrubs--their biology and utilization: An international symposium: Proceedings; 1971 July; Logan, UT. Gen. Tech. Rep. INT-1. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 101-108. [22754]
95. Kamill, Bernardette W.; Steinberger, Y.; Whitford, W. G. 1985. Soil microarthropods from the Chihuahuan Desert of New Mexico. Journal of Zoology. 205(2): 273-286. [11560]
96. Kartesz, J. T. The Biota of North America Program (BONAP). 2015. North American Plant Atlas, [Online]. Chapel Hill, NC: The Biota of North America Program (Producer). Available: http://bonap.net/napa [maps generated from Kartesz, J. T. 2015. Floristic synthesis of North America, Version 1.0. Biota of North America Program (BONAP)] (in press). [94573]
97. Kartesz, J. T., The Biota of North America Program (BONAP). 2015. Taxonomic Data Center, [Online]. Chapel Hill, NC: The Biota of North America Program (Producer). Available: http://bonap.net/tdc [maps generated from Kartesz, J. T. 2015. Floristic synthesis of North America, Version 1.0. Biota of North America Program (BONAP)] (in press). [84789]
98. Kartesz, John Thomas. 1988. A flora of Nevada. Reno, NV: University of Nevada. 1729 p. Dissertation. [42426]
99. Keane, Robert E.; Mincemoyer, Scott A.; Schmidt, Kirsten M.; Long, Donald G.; Garner, Janice L. 2000. Mapping vegetation and fuels for fire management on the Gila National Forest Complex, New Mexico. Gen. Tech. Rep. RMRS-GTR-46. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 126 p. [91538]
100. Kearney, Thomas H.; Peebles, Robert H.; Howell, John Thomas; McClintock, Elizabeth. 1960. Arizona flora. 2nd ed. Berkeley, CA: University of California Press. 1085 p. [6563]
101. Kehr, Jacqueline M.; Merritt, David M.; Stromberg, Juliet C. 2014. Linkages between primary seed dispersal, hydrochory and flood timing in a semi-arid region river. Journal of Vegetation Science. 25(1): 287-300. [95960]
102. Killingbeck, K. T.; Whitford, W. G. 2001. Nutrient resorption in shrubs growing by design, and by default in Chihuahuan Desert arroyos. Oecologia. 128(3): 351-359. [78930]
103. Kittams, Walter H.; Evans, Stanley L.; Cooke, Derrick C. 1979. Food habits of mule deer on foothills of Carlsbad Caverns National Park. In: Genoways, Hugh H.; Baker, Robert J., eds. Biological investigations in the Guadalupe Mountains National Park: Proceedings of a symposium; 1975 April 4-5; Lubbock, TX. Proceedings and Transactions Series No. 4. Washington, DC: U.S. Department of the Interior, National Park Service: 403-426. [16023]
104. Kozma, Jeffrey M.; Mathews, Nancy E. 1997. Breeding bird communities and nest plant selection in Chihuahuan Desert habitats in south-central New Mexico. The Wilson Bulletin. 109(3): 424-436. [78942]
105. Kraus, Helen Tyler. 1998. Effects of mulch on soil moisture and growth of desert willow. HortTechnology. 8(4): 588-590. [95962]
106. Krausman, Paul R.; Kuenzi, Amy J.; Etchberger, Richard C.; Rautenstrauch, Kurt T.; Ordway, Leonard L.; Hervert, John J. 1997. Diets of mule deer. Journal of Range Management. 50(5): 513-522. [27845]
107. Kuchler, A. W. 1964. Manual to accompany the map of potential vegetation of the conterminous United States. Special Publication No. 36. New York: American Geographical Society. 166 p. [1384]
108. Lamb, S. H. 1971. Woody plants of New Mexico and their value to wildlife. Bulletin No. 14. Albuquerque, NM: New Mexico Department of Game and Fish. 80 p. [9818]
109. Lambert, Adam M.; D'Antonio, Carla M.; Dudley, Tom L. 2010. Invasive species and fire in California ecosystems. Fremontia. 38(2): 29-36. [89006]
110. LANDFIRE Biophysical Settings. 2009. Biophysical setting 1511552: North American warm desert riparian systems - Stringers. In: LANDFIRE Biophysical Setting Model: Map zone 15, [Online]. Washington, DC: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory; U.S. Geological Survey; Arlington, VA: The Nature Conservancy (Producers). Available: https://www.fs.usda.gov/database/feis/pdfs/BpS/1511552.pdf. [96194]
111. LANDFIRE. 2008. CONUS refresh (LANDFIRE 1.1.0). Biophysical settings layer, LANDFIRE data distribution site, [Online]. In: LANDFIRE. U.S. Department of the Interior, Geological Survey (Producer). Available: https://landfire.cr.usgs.gov/viewer/ [2017, January 10]. [89416]
112. Latta, Marjorie J.; Beardmore, Carol J.; Corman, Troy E. 1999. Arizona Partners in Flight bird conservation plan, [Version 1.0]. Technical Report 142. Phoenix, AZ: Arizona Game and Fish Department, Nongame and Endangered Wildlife Program. 334 p. Available: https://www.partnersinflight.org/wp-content/uploads/2017/03/Arizona-State-Plan-v-1-1999-1.pdf [2021, December 15]. [63869]
113. Lei, Simon A.; Walker, Lawrence R. 1997. Classification and ordination of Coleogyne communities in southern Nevada. The Great Basin Naturalist. 57(2): 155-162. [27721]
114. Lei, Steven A.; Lei, Simon A. 1999. Ecology of psammophytic plants in the Mojave, Sonoran, and Great Basin Deserts. In: McArthur, E. Durant; Ostler, W. Kent; Wambolt, Carl L., compilers. Proceedings: Shrubland ecotones; 1998 August 12-14; Ephraim, UT. Proceedings RMRS-P-11. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 212--216. [36089]
115. Leiberg, John B. 1900. The San Jacinto Forest Reserve. In: Walcott, C. D.; Gannett, Henry. Twentieth annual report of the United States Geological Survey to the Secretary of the Interior, 1898-1899. Washington, DC: U.S. Government Printing Office: 455-478. [90777]
116. Levin, Geoffrey A. 1988. How plants survive in the desert. Environment Southwest. Summer: 20-25. [9239]
117. Levine, C. M.; Stromberg, J. C. 2001. Effects of flooding on native and exotic plant seedlings: Implications for restoring south-western riparian forests by manipulating water and sediment flows. Journal of Arid Environments. 49(1): 111-131. [43624]
118. Lite, S. J.; Stromberg, J. C. 2005. Surface water and ground-water thresholds for maintaining Populus-Salix forests, San Pedro River, Arizona. Biological Conservation. 125(2): 153-167. [53987]
119. Little, Elbert L., Jr. 1950. Southwestern trees: A guide to the native species of New Mexico and Arizona. Agric. Handb. No. 9. Washington, DC: U.S. Department of Agriculture, Forest Service. 109 p. [20317]
120. Little, Elbert L., Jr. 1976. Atlas of United States trees. Volume 3. Minor western hardwoods. Misc. Publ. 1314. Washington, DC: U.S. Department of Agriculture, Forest Service. 13 p. [+ 290 maps]. [10430]
121. Little, Elbert L., Jr. 1979. Checklist of United States trees (native and naturalized). Agric. Handb. 541. Washington, DC: U.S. Department of Agriculture, Forest Service. 375 p. [2952]
122. Love, Stephen L.; Akins, Candace J. 2019. Second summary of the native seed germination studies of Norman C Deno: Species with names beginning with letters C through E. Native Plants Journal. 20(1): 65-97. [95898]
123. Lovich, Jeffrey E.; Egan, Thomas B.; de Gouvenain, Roland C. 1994. Tamarisk control on public lands in the desert of southern California: Two case studies. In: Environmental stewardship through weed control: Proceedings, 46th annual California Weed Science Society conference; 1994 January 17-19; San Jose, California. No. 46. Fremont, CA: California Weed Science Society: 166-177. [44086]
124. Lowe, Charles H., Jr. 1961. Biotic communities in the sub-Mogollon region of the Inland Southwest. Arizona Academy of Science Journal. 2: 40-49. [20379]
125. MacMahon, James A. 1988. Warm deserts. In: Barbour, Michael G.; Billings, William Dwight, eds. North American terrestrial vegetation. New York: Cambridge University Press: 231-264. [19547]
126. MacMahon, James A.; Schimpf, David J. 1981. Water as a factor in the biology of North American desert plants. In: Evans, D. D.; Thames, J. L., eds. Water in desert ecosystems. Stroudsburg, PA: Dowden, Hutchinson, and Ross: 114-171. [30888]
127. Magill, Arthur M.; Miller, Carol; Rodgers, Jane E. 2008. Chilopsis linearis (Cov.) Sweet. In: Bonner, Franklin T.; Karrfalt, Robert P., eds. Woody plant seed manual. Agric. Handbook No. 727. Washington, DC: U.S. Department of Agriculture, Forest Service: 396-397. [79134]
128. Magill, Arthur W. 1974. Chilopsis linearis (Cav.) Sweet desert willow. In: Schopmeyer, C. S., technical coordinator. Seeds of woody plants in the United States. Agric. Handb. 450. Washington, DC: U.S. Department of Agriculture, Forest Service: 321-322. [7587]
129. Martin, William C.; Hutchins, Charles R. 1981. A flora of New Mexico. Volume 2. Germany: J. Cramer. 2589 p. [37176]
130. Mason, Charles T.; Mason, Patricia B. 1987. A handbook of Mexican roadside flora. 1st ed. Tucson, AZ: The University of Arizona Press. 380 p. [93925]
131. McClaran, Mitchel P.; Brady, Ward W. 1994. Arizona's diverse vegetation and contributions to plant ecology. Rangelands. 16(5): 208-217. [29721]
132. Miksicek, Charles H. 1983. Appendix B: Plant remains from agricultural features. In: Teague, Lynn S.; Crown, Patricia L., eds. Hohokam archaeology along the Salt-Gila Aqueduct: Central Arizona Project. Archaeological Series No. 150. Tucson, AZ: Arizona State Museum, Cultural Resource Management Section: 604-620. [Vol. 3: Environment and subsistence; Bureau of Reclamation Contract No. 0-07-32-V0101]. [44341]
133. Miksicek, Charles H. 1984. Historic desertification, prehistoric vegetation change, and Hohokam subsistence in the Salt-Gila Basin. In: Teague, Lynn S.; Crown, Patricia L., eds. Hohokam archaeology along the Salt-Gila Aqueduct: Central Arizona Project. Archaeological Series No. 150. Tucson, AZ: Arizona State Museum, Cultural Resource Management Section: 53-80. [Vol. 7: Environment and subsistence; Bureau of Reclamation Contract No. 0-07-32-V0101]. [44339]
134. Milchunas, Daniel G., ed. 2006. Responses of plant communities to grazing in the southwestern United States. Gen. Tech. Rep. RMRS-GTR-169. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 126 p. [63342]
135. Milton, Suzanne J.; Dean, W. R. J.; Kerley, G. I. H.; Hoffman, M. T.; Whitford, W. G. 1998. Dispersal of seeds as nest material by the cactus wren. The Southwestern Naturalist. 43(4): 449-452. [29454]
136. Minckley, W. L.; Brown, David E. 1982. Wetlands. In: Brown, David E., ed. Biotic communities of the American Southwest--United States and Mexico. Desert Plants. 4(1-4): 223-287. [8898]
137. Minckley, W. L.; Clark, Thomas O. 1981. Vegetation of the Gila River Resource Area, eastern Arizona. Desert Plants. 3(3): 124-140. [10863]
138. Moore, Michael. 1989. Medicinal plants of the desert and canyon West. Santa Fe, NM: Museum of New Mexico Press. 184 p. [25027]
139. Morton, Howard L. 1994. SRM 507: Palo verde - cactus. In: Shiflet, Thomas N., ed. Rangeland cover types of the United States. Denver, CO: Society for Range Management: 66-67. [67047]
140. Muldavin, Esteban H.; DeVelice, Robert L.; Ronco, Frank, Jr. 1996. A classification of forest habitat types: Southern Arizona and portions of the Colorado Plateau. RM-GTR-287. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 130 p. [27968]
141. Muldavin, Esteban; Durkin, Paula; Bradley, Mike; Stuever, Mary; Mehlhop, Patricia. 2000. Handbook of wetland vegetation communities of New Mexico. Volume 1: Classification and community descriptions. Albuquerque, NM: University of New Mexico, Biology Department; New Mexico Natural Heritage Program. 172 p. [+ appendices]. [45517]
142. Nagler, Pamela L.; Glenn, Edward P.; Jarnevich, Catherine S. J.; Shafroth, Patrick B. 2011. Distribution and abundance of saltcedar and Russian olive in the western United States. Critical Reviews in Plant Sciences. 30(6): 508–523. [96197]
143. Naiman, Robert J.; Decamps, Henri; Pollock, Michael. 1993. The role of riparian corridors in maintaining regional biodiversity. Ecological Applications. 3(2): 209-212. [96206]
144. NatureServe. 2009. International ecological classification standard: Terrestrial ecological classifications. In: NatureServe Central Databases. Arlington, VA: NatureServe (Producer). 1172 p. Available: http://downloads.natureserve.org/get_data/data_sets/veg_data/nsDescriptions.pdf [2020, August 3]. [94380]
145. NatureServe. 2018. International ecological classification standards: Terrestrial ecological classifications. NVC groups of the eastern U.S. on the LANDFIRE legend. Arlington, VA: NatureServe. 696 p. Available: https://landfire.gov/documents/LANDFIRE_Natural_NVC_Groups_Descriptions_EasternUS.pdf [2021, February 18]. [95146]
146. Nevada Department of Conservation and Natural Resources, Nevada Natural Heritage Program. 2003. National vegetation classification for Nevada, [Online]. Carson City, NV: Nevada Department of Conservation and Natural Resources, Nevada Natural Heritage Program (Producer). 15 p. Available: https://heritage.nv.gov/documents/national-vegetation-classification-nvc--info-from-usgs [2021, December 6]. [55021]
147. Newman, George A. 1979. Compositional aspects of breeding avifaunas in selected woodlands of the southern Guadalupe Mountains, Texas. In: Genoways, Hugh H.; Baker, Robert J., eds. Biological investigations in the Guadalupe Mountains National Park: Proceedings of a symposium; 1975 April 4-5; Lubbock, TX. Proceedings and Transactions Series No. 4. Washington, DC: U.S. Department of the Interior, National Park Service: 181-237. [16021]
148. Nichol, A. A. 1952. [Revised]. The natural vegetation of Arizona. Tech. Bull. 68. Tucson, AZ: University of Arizona, Agricultural Experiment Station: 189-230. [Revisions by W. S. Phillips]. [3928]
149. Nilsen, Erik Tallak; Sharifi, M. Rasoul; Rundel, Philip W. 1984. Comparative water relations of phreatophytes in the Sonoran Desert of California. Ecology. 65(3): 767-778. [49012]
150. Odening, Walter R.; Strain, B. R.; Oechel, W. C. 1974. The effect of decreasing water potential on net CO2 exchange of intact desert shrubs. Ecology. 55(5): 1086-1095. [17775]
151. O’Connor, Christopher D.; Falk, Donald A.; Garfin, Gregg M. 2020. Projected climate-fire interactions drive forest to shrubland transition on an Arizona Sky Island. Frontiers in Environmental Science. 8(137): 1-17. [95923]
152. Pace, Marcelo R.; Angyalossy, Veronica. 2013. Wood anatomy and evolution: A case study in the Bignoniaceae. International Journal of Plant Sciences. 172(7): 1014-1048. [95972]
153. Pace, Marcelo R.; Lohmann, Lucia G.; Olmstead, Richard G.; Angyalossy, Veronica. 2015. Wood anatomy of major Bignoniaceae clades. Plant Systematics and Evolution. 301(3): 967-995. [95974]
154. Pase, Charles P.; Layser, Earle F. 1977. Classification of riparian habitat in the Southwest. In: Johnson, Roy; Jones, Dale A., technical coordinators. Importance, preservation and management of riparian habitat: A symposium; 1977 July 9; Tucson, AZ. Gen. Tech. Rep. RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 5-9. Available from: NTIS, Springfield, VA 22151; PB-274 582. [5333]
155. Paysen, Timothy E.; Derby, Jeanine A.; Black, Hugh, Jr.; Bleich, Vernon C.; Mincks, John W. 1980. A vegetation classification system applied to southern California. Gen. Tech. Rep. PSW-45. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 33 p. [1849]
156. Peinado, M.; Macias, M. A.; Ocana-Peinado, F. M.; Aguirre, J. L.; Delgadillo, J. 2011. Bioclimates and vegetation along the Pacific basin of Northwestern Mexico. Plant Ecology. 212(2): 263-281. [95975]
157. Pemberton, Robert W. 1988. The abundance of plants bearing extrafloral nectaries in Colorado and Mojave Desert communities of southern California. Madrono. 35(3): 238-246. [6163]
158. Petersen, C.; Brown, J. H.; Kodric-Brown, A. 1982. An experimental study of floral display and fruit set in Chilopsis linearis (Bignoniaceae). Oecologia. 55(1): 7-11. [12061]
159. Peterson, Eric B. 2008. International vegetation classification alliances and associations occurring in Nevada with proposed additions. Carson City, NV: Nevada Natural Heritage Program. 347 p. [77864]
160. Pettit, Neil E.; Naiman, Robert J. 2007. Fire in the riparian zone: Characteristics and ecological consequences. Ecosystems. 10(5): 673-687. [68289]
161. Powell, A. Michael. 1988. Trees and shrubs of Trans-Pecos Texas: Including Big Bend and Guadalupe Mountains National Parks. Big Bend National Park, TX: Big Bend Natural History Association. 536 p. [6130]
162. Price, Theresa Lorez. 2008. Flora of Mount Ord, central Arizona and habitat characteristics of Fremontodendron californicum. Tempe, AZ: Arizona State University. 120 p. Thesis. [94820]
163. Racher, Brent J.; Britton, Carlton M. 2003. Fire in saltcedar ecosystems. In: Proceedings: Saltcedar and water resources symposium; 2003 July 16–17; San Angelo, TX. [Publisher name unknown] 94–99. [96196]
164. Raunkiaer, C. 1934. The life forms of plants and statistical plant geography. Oxford, England: Clarendon Press. 632 p. [2843]
165. Read, Ralph A.; Sprackling, John. 1980. Cottonwood-willow--Forest Cover Type 235. In: Eyre, F. H., ed. Forest cover types of the United States and Canada. Washington, DC: Society of American Foresters: 113 p. [50049]
166. Reeves, Timothy. 1976. Vegetation and flora of Chiricahua National Monument, Cochise County, Arizona. Tempe, AZ: Arizona State University. 180 p. Thesis. [20385]
167. Reina-Guerrero, Ana Lilia; Van Devender, Thomas R. 2013. Chihuahuan Desert flora of La Calera, Municipio de Agua Prieta, Sonora, Mexico. In: Gottfried, Gerald J.; Ffolliott, Peter F.; Gebow, Brooke S.; Eskew, Lane G.; Collins, Loa C., comps. Proceedings: Merging science and management in a rapidly changing world: Biodiversity and management of the Madrean Archipelago III. RMRS-P-67; 2012 May 1-5; Tucson, AZ. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 225-228. [96217]
168. Reynolds, Hudson G.; Johnson, R. Roy. 1964. Habitat relations of vertebrates of the Sierra Ancha Experimental Forest. Res. Pap. RM-4. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 16 p. [13485]
169. Reynolds, Lindsay V.; Cooper, David J. 2011. Ecosystem response to removal of exotic riparian shrubs and a transition to upland vegetation. Plant Ecology. 212(8): 1243-1261. [83418]
170. Rice, Peter M.; McPherson, Guy R.; Rew, Lisa J. 2008. Fire and nonnative invasive plants in the Interior West bioregion. In: Zouhar, Kristin; Smith, Jane Kapler; Sutherland, Steve; Brooks, Matthew L., eds. Wildland fire in ecosystems: Fire and nonnative invasive plants. Gen. Tech. Rep. RMRS-GTR-42-vol. 6. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 141-173. [70332]
171. Richardson, Sarah C. 2004. Are nectar-robbers mutualists or antagonists? Oecologia. 139(2): 246-254. [95978]
172. Richardson, Sarah C. 2004. Benefits and costs of floral visitors to Chilopsis linearis: Pollen deposition and stigma closure. Oikos. 107(2): 363-375. [95977]
173. Richardson, W. Scott; Cartron, Jean-Luc E.; Krueper, David J.; Turner, Lauren; Skinner, Thomas H. 2000. The status of the cactus ferruginous pygmy-owl in Arizona: Population surveys and habitat assessment. In: Cartron, Jean-Luc E.; Finch, Deborah M., tech. eds. Ecology and conservation of the cactus ferruginous pygmy-owl in Arizona. Gen. Tech. Rep. RMRS-GTR-43. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 27-39. [35491]
174. Ringold, Paul L.; Magee, Teresa K.; Peck, David V. 2008. Twelve invasive plant taxa in US western riparian ecosystems. Journal of the North American Benthological Society. 27(4): 949-966. [73064]
175. Rodriguez, Elena; Parsons, Jason G.; Peralta-Videa, Jose R.; Cruz-Jimenez, Gustavo; Romero-Gonzalez, Jaime; Sanchez-Salcido, Blanca E.; Saupe, Geoffrey B.; Duarte-Gardea, Maria; Gardea-Torresdey, Jorge L. 2007. Potential of Chilopsis linearis for gold phytomining: Using Xas to determine gold reduction and nanoparticle formation within plant tissues. International Journal of Phytoremediation. 9(2): 133-147. [95980]
176. Rucks, Michael G. 1984. Composition and trend of riparian vegetation on five perennial streams in southeastern Arizona. In: Warner, Richard E.; Hendrix, Kathleen M., eds. California riparian systems: Ecology, conservation, and productive management: Proceedings of a conference; 1981 September 17-19; Davis, CA. Berkeley, CA: University of California Press: 97-107. [5831]
177. Rundel, Philip W.; Gibson, Arthur C. 1996. Ecological communities and processes in a Mojave Desert ecosystem: Rock Valley, Nevada. Cambridge; New York: Cambridge University Press. 369 p. [61799]
178. Sadoti, Giancarlo. 2008. Nest-site selection by common black-hawks in southwestern New Mexico. Journal of Field Ornithology. 79(1): 11-19. [96285]
179. Schaefer, Douglas; Steinberger, Yosef; Whitford, Walter G. 1985. The failure of nitrogen and lignin control of decomposition in a North American desert. Oecologia. 65(3): 382-386. [95981]
180. Scott, F. Murray. 1935. Contribution to the causal anatomy of the desert willow, Chilopsis linearis. American Journal of Botany. 22(3): 333-343. [95982]
181. Seavy, Nathaniel E.; Gardali, Thomas; Golet, Gregory H.; Griggs, F. Thomas; Howell, Christine A.; Kelsey, Rodd; Small, Stacy L.; Viers, Joshua H.; Weigand, James F. 2009. Why climate change makes riparian restoration more important than ever: Recommendations for practice and research. Ecological Restoration. 27(3): 330-338. [96229]
182. Shafroth, Patrick B.; Beauchamp, Vanessa B.; Briggs, Mark K.; Lair, Kenneth; Scott, Michael L.; Sher, Anna A. 2008. Planning riparian restoration in the context of tamarix control in western North America. Restoration Ecology. 16(1): 97-112. [74711]
183. Sharifi, M. R.; Meinzer, F. C.; Nilsen, E. T.; Rundel, P. W.; Virginia, R. A.; Jarrell, W. M.; Herman, D. J.; Clark, P. C. 1988. Effect of manipulation of water and nitrogen supplies on the quantitative phenology of Larrea tridentata (creosote bush) in the Sonoran Desert. American Journal of Botany. 75(8): 1163-1174. [5476]
184. Sharifi, M. R.; Meinzer, F. C.; Rundel, P. W.; Nilsen, E. T. 1990. Effect of manipulating soil water and nitrogen regimes on clipping production and water relations of creosote bush. In: McArthur, E. Durant; Romney, Evan M.; Smith, Stanley D.; Tueller, Paul T., compilers. Proceedings--symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management; 1989 April 5-7; Las Vegas, NV. Gen. Tech. Rep. INT-276. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 245-249. [12857]
185. Shiflet, Thomas N., ed. 1994. Rangeland cover types of the United States. Denver, CO: Society for Range Management. 152 p. [23362]
186. Short, Henry L. 1977. Food habits of mule deer in a semi-desert grass-shrub habitat. Journal of Range Management. 30(3): 206-209. [9895]
187. Sidman, Gabriel; Guertin, D. Phillip; Goodrich, David C.; Thoma, David; Falk, Donald; Burns, I. Shea. 2016. A coupled modelling approach to assess the effect of fuel treatments on post-wildfire runoff and erosion. International Journal of Wildland Fire. 25(3): 351-362. [96201]
188. Simpson, Benny J. 1988. A field guide to Texas trees. Austin, TX: Texas Monthly Press. 372 p. [11708]
189. Simpson, Benny J.; Hipp, Billy W.; McWilliams, Edward L. 1989. 'White Storm' and 'Dark Storm' desert willow. HortScience. 24(1): 178-179. [12244]
190. Skartvedt, Peter H. 2000. Woody riparian vegetation patterns in the upper Mimbres Watershed, southwestern New Mexico. The Southwestern Naturalist. 45(1): 6-14. [38910]
191. Smith, D. Max; Finch, Deborah M. 2016. Riparian trees and aridland streams of the southwestern United States: An assessment of the past, present, and future. Journal of Arid Environments. 135(23): 120-131. [96185]
192. Smith, D. Max; Finch, Deborah M. 2017. Climate change and wildfire effects in aridland riparian ecosystems: An examination of current and future conditions. Gen. Tech. Rep. RMRS-GTR-364. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 65 p. [96256]
193. Smith, D. Max; Finch, Deborah M.; Gunning, Christian; Jamison, Roy; Kelly, Jeffrey F. 2009. Post-wildfire recovery of riparian vegetation during a period of water scarcity in the southwestern USA. Fire Ecology--Special Issue. 5(1): 38-55. [77004]
194. Smith, G. Shannon; Pittcock, Kim. 1989. The collector's quest. American Nurseryman. 169(1): 56-65. [12243]
195. Steger, Robert E.; Beck, Reldon F. 1973. Range plants as ornamentals. Journal of Range Management. 26(1): 72-74. [12038]
196. Stevens, Larry E. 2002. Exotic tamarisk on the Colorado Plateau, [Online]. In: Grahame, John D.; Sisk, Thomas D., eds. Canyons, cultures and environmental change: An introduction to the land-use history of the Colorado Plateau. U.S. Geological Survey (Producer). Available: https://web.archive.org/web/20060206062038/http://www.cpluhna.nau.edu/Biota/tamarisk.htm [2021, December 15, 2021]. [44465]
197. Stevens, Lawrence E.; Brown, Bryan T.; Simpson, James M.; Johnson, R. Roy. 1977. The importance of riparian habitat to migrating birds. In: Johnson, Raymond Roy; Jones, Dale A., tech. coords. Symposium on the importance, preservation and management of riparian habitat; 1977 July 9; Tucson, AZ. GTR RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 156-164. [85073]
198. Stickney, Peter F. 1989. Seral origin of species comprising secondary plant succession in northern Rocky Mountain forests. FEIS workshop: Postfire regeneration. Unpublished draft on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Missoula Fire Sciences Laboratory. 10 p. [20090]
199. Strain, B. R.; Chase, Valerie C. 1966. Effect of past an prevailing temperatures on the carbon dioxide exchange capacities of some woody desert perennials. Ecology. 47(6): 1043-1045. [95983]
200. Stromberg, J. C. 1993. Fremont cottonwood-Goodding willow riparian forests: A review of their ecology, threats, and recovery potential. Journal of the Arizona-Nevada Academy of Sciences. 27(1): 97-110. [29724]
201. Stromberg, J. C.; McCluney, K. E.; Dixon, M. D.; Meixner, T. 2013. Dryland riparian ecosystems in the American Southwest: Sensitivity and resilience to climatic extremes. Ecosystems. 16(3): 411-415. [96205]
202. Stromberg, J. C.; Tiller, R.; Richter, B. 1996. Effects of groundwater decline on riparian vegetation of semiarid regions: The San Pedro, Arizona. Ecological Applications. 6(1): 113-131. [48991]
203. Stromberg, Juliet C. 1998. Functional equivalency of saltcedar (Tamarix chinensis) and Fremont cottonwood (Populus fremontii) along a free-flowing river. Wetlands. 18(4): 675-686. [43989]
204. Stromberg, Juliet C.; Rychener, Tyler J. 2010. Effects of fire on riparian forests along a free-flowing dryland river. Wetlands. 30(1): 75-86. [81928]
205. Strong, Thomas R., Bock, Carl E. 1990. Bird species distribution patterns in riparian habitats in southeastern Arizona. The Condor. 92(4): 866-885. [69088]
206. Stuever, Mary C. 1997. Fire-induced mortality of Rio Grande cottonwood. Albuquerque, NM: University of New Mexico. 85 p. Thesis. [44706]
207. Stuever, Mary C.; Hayden, John S. 1996. Plant associations (habitat types) of the forests and woodlands of Arizona and New Mexico. Final report: Contract R3-95-27. Placitas, NM: Seldom Seen Expeditions. 520 p. [28868]
208. Szaro, Robert C. 1989. Riparian forest and scrubland community types of Arizona and New Mexico. Desert Plants. 9(3-4): 70-138. [604]
209. Szaro, Robert C.; Pase, Charles P. 1983. Short-term changes in a cottonwood-ash-willow association on a grazed and an ungrazed portion of Little Ash Creek in central Arizona. Journal of Range Management. 36(3): 382-384. [6520]
210. Temple, Patrick J. 1989. Oxidant air pollution effects on plants of Joshua Tree National Monument. Environmental Pollution. 57(1): 35-47. [47314]
211. Texas Natural Heritage Program. 1993. Plant communities of Texas (Series level). Austin, TX: Texas Parks and Wildlife Department. Unpublished report on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 26 p. [23810]
212. Thomas, Jonathan A.; White, Joseph D.; Murray, Darrel B. 2016. Tree species influence woodland canopy characteristics and crown fire potential. Forest Ecology and Management. 362: 169-176. [94522]
213. Thompson, Robert S.; Anderson, Katherine H.; Bartlein, Patrick J. 1999. Digital representations of tree species range maps from "Atlas of United States trees" by Elbert L. Little, Jr. (and other publications). In: Atlas of relations between climatic parameters and distributions of important trees and shrubs in North America. Denver, CO: U.S. Geological Survey, Information Services (Producer). On file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT; FEIS files. [92575]
214. Thorne, Robert F. 1976. The vascular plant communities of California. In: Latting, June, ed. Symposium proceedings: Plant communities of southern California; 1974 May 4; Fullerton, CA. Special Publication No. 2. Berkeley, CA: California Native Plant Society: 1-31. [3289]
215. Thorne, Robert F. 1982. The desert and other transmontane plant communities of southern California. Aliso. 10(2): 219-257. [3768]
216. Tipton, Jimmy L. 1988. 'Marfa Lace', 'Alpine', and 'Tejas' desert willows. HortScience. 23(4): 782. [12065]
217. Tipton, Jimmy L. 1988. Drought resistance of Chilopsis linearis, Morus alba, and Tecoma stans. HortScience. 23(3): 721. Abstract. [95984]
218. Tratz, Wallace M.; Vogl, Richard J. 1977. Postfire vegetational recovery, productivity, and herbivore utilization of a chaparral-desert ecotone. In: Mooney, Harold A.; Conrad, C. Eugene, technical coordinators. Proceedings of the symposium on the environmental consequences of fire and fuel management in Mediterranean ecosystems; 1977 August 1-5; Palo Alto, CA. Gen. Tech. Rep. WO-3. Washington, DC: U.S. Department of Agriculture, Forest Service: 426-430. [4873]
219. Tratz, Wallace Michael. 1978. Postfire vegetational recovery, productivity, and herbivore utilization of a chaparral-desert ecotone. Los Angeles, CA: California State University. 133 p. Thesis. [5495]
220. Turner, Raymond M.; Bowers, Janice E.; Burgess, Tony L. 2005. Sonoran Desert plants: An ecological atlas. Tucson, AZ: The University of Arizona Press. 504 p. [94363]
221. Turner, Raymond M.; Webb, Robert H.; Esque, Todd C.; Rogers, Garry F. 2010. Repeat photography and low-elevation fire responses in the southwestern United States. In: Webb, Robert H.; Boyer, Diane E.; Turner, Raymond M. Repeat photography: Methods and applications in the natural sciences. 1st ed. Washington, DC: Island Press: 224-235. [95737]
222. U.S. Fish and Wildlife Service. 2022. Listed species believed or known to occur in New Mexico, [Online]. Environmental Conservation Online System. Available: https://ecos.fws.gov/ecp0/reports/species-listed-by-state-report?state=NM [2022, January 2]. [96188]
223. USDA, Natural Resources Conservation Service. 1997. Riparian forest buffer. Conservation Practice Job Sheet 391. U.S. Department of Agriculture, Natural Resources Conservation Service. 4 p. [96191]
224. USDA, NRCS. 2021. The PLANTS Database, [Online]. Greensboro, NC: U.S. Department of Agriculture, Natural Resources Conservation Service, National Plant Data Team (Producer). Available: https://plants.usda.gov/. [34262]
225. USDI, Fish and Wildlife Service. 2022. Endangered Species Program, [Online]. U.S. Department of the Interior, Fish and Wildlife Service (Producer). Available: https://www.fws.gov/endangered/. [86564]
226. Van Buren, Renee; Cooper, Janet G.; Shultz, Leila M.; Harper, Kimball T. 2011. Woody plants of Utah. A field guide with identification keys to native and naturalized trees, shrubs, cacti, and vines. Logan, UT: Utah State University Press. 513 p. [93924]
227. Van Dersal, William R. 1938. Native woody plants of the United States, their erosion-control and wildlife values. Misc. Publ. No. 303. Washington, DC: U.S. Department of Agriculture. 362 p. [4240]
228. Van Nguyen, Anh; Deineka, Victor Ivanovic; Deineka, Lyudmila Aleksandrovna; Vu, Anh Thi Ngoc. 2019. Determination of triacylglycerols and fatty acid composition of Chilopsis linearis seed oil by reverse-phase high-performance liquid chromatography. Turkish Journal of Chemistry. 43(3): 881-889. [95971]
229. van Riper, Charles III; Paxton, Kristina L.; O’Brien, Chris; Shafroth, Patrick B.; McGrath, Laura J. 2008. Rethinking avian response to Tamarix on the Lower Colorado River: A threshold hypothesis. Restoration Ecology. 16(1): 155 167. [96203]
230. Vega-Mares, Jose Humberto; Estrada-Castillon, Andres Eduardo; Villarreal-Quintanilla, Jose Angel; Martinez, Gustavo Quintana. 2014. Flora of the halophytic grasslands in the Valle de Janos, Chihuahua, Mexico. Journal of the Botanical Research Institute of Texas. 8(1): 151-163. [95985]
231. Villar, R.; Merino, J. 2001. Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems. New Phytologist. 151(1): 213-226. [80064]
232. Vines, Robert A. 1960. Trees, shrubs, and woody vines of the Southwest. Austin, TX: University of Texas Press. 1104 p. [7707]
233. Vogl, Richard J. 1976. An introduction to the plant communities of the Santa Ana and San Jacinto Mountains. In: Latting, June, ed. Symposium proceedings: Plant communities of southern California; 1974 May 4; Fullerton, CA. Special Publication No. 2. Berkeley, CA: California Native Plant Society: 77-98. [4230]
234. Walker, Lawrence R.; Barnes, Pamela L.; Powell, Elizabeth A. 2006. Tamarix aphylla: A newly invasive tree in southern Nevada. Western North American Naturalist. 66(2): 191-201. [64514]
235. Wallmo, O. C. 1955. Vegetation of the Huachuca Mountains, Arizona. The American Midland Naturalist. 54(2): 466-480. [20325]
236. Walters, M. Alice; Teskey, Robert O.; Hinckley, Thomas M. 1980. Impact of water level changes on woody riparian and wetland communities. Volume 7: Mediterranean region, western arid and semi-arid region. FWS/OBS-78/93. Washington, DC: U.S. Department of the Interior, Fish and Wildlife Service, Biological Services Program. 84 p. [52899]
237. Walters, R. S.; Auchmoody, L. R. 1989. Vegetation re-establishment on a hardwood forest site denuded by brine. Landscape and Urban Planning. 17(2): 127-133. [9819]
238. Warnock, Barton. 1994. SRM 713: Grama-muhly threeawn. In: Shiflet, Thomas N., ed. Rangeland cover types of the United States. Denver, CO: Society for Range Management: 92-93. [67363]
239. Warren, Peter L.; Hoy, Marina S.; Hoy, Wilton E. 1992. Vegetation and flora of Fort Bowie National Historic Site, Arizona. Tech. Rep. NPS/WRUA/NRTR-92/43. Tucson, AZ: The University of Arizona, School of Renewable Natural Resources, Cooperative National Park Resources Studies Unit. 78 p. [19871]
240. Wasser, Clinton H. 1982. Ecology and culture of selected species useful in revegetating disturbed lands in the West. FWS/OBS-82/56. Washington, DC: U.S. Department of the Interior, Fish and Wildlife Service, Office of Biological Services, Western Energy and Land Use Team. 347 p. Available from NTIS, Springfield, VA 22161; PB-83-167023. [2458]
241. Wauer, Roland H. 1971. Ecological distribution of birds of the Chisos Mountains, Texas. The Southwestern Naturalist. 16(1): 1-29. [24969]
242. Wauer, Roland H. 1977. Significance of Rio Grande riparian systems upon the avifauna. In: Johnson, R. Roy; Jones, Dale A., tech. coords. Importance, preservation and management of riparian habitat: A symposium; 1977 July 9; Tucson, AZ. Gen. Tech. Rep. RM-43. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 165-174. [5344]
243. Webb, Amanda D. 2017. Fire effects and management in riparian ecosystems of the southwestern United States and Mexico. Tucson, AZ: University of Arizona. 168 p. Thesis. [92607]
244. Webb, Amanda D.; Falk, Donald A.; Finch, Deborah M. 2019. Fire ecology and management in lowland riparian ecosystems of the southwestern United States and northern Mexico. Gen. Tech. Rep. RMRS-GTR-401. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 132 p. [95328]
245. Webb, Robert H.; Griffiths, Peter G.; Wallace, Cynthia S. A.; Boyer, Diane E. 2007. Channel response to low-elevation desert fire: The King Valley Fire of 2005. Rep. Series Data–DS 275. Tucson, AZ: U.S. Department of the Interior. U.S. Geological Survey. 52 p. [95986]
246. Went, F. W. 1948. Ecology of desert plants. I. Observations on germination in the Joshua Tree National Monument, California. Ecology. 29(3): 242-253. [12915]
247. Whitfield, Charles J.; Anderson, Hugh L. 1938. Secondary succession in the desert plains grassland. Ecology. 19(2): 171-180. [5252]
248. Whitham, Thomas G. 1977. Coevolution of foraging in Bombus and nectar dispensing in Chilopsis: A last dreg theory. Science. 197(4303): 593-596. [96218]
249. Wickersham, Lynn E.; Johnson, Kristine; Sadoti, Giancarlo; Neville, Teri; Wickersham, John. 2020. Habitat use at multiple scales by nesting gray vireos in New Mexico. In: Malcolm, Karl; Dykstra, Brian; Johnson, Kristine; Lightfoot, David; Muldavin, Esteban; Ramsey, Marikay, eds. Symposium proceedings on pinon-juniper habitats: Status and management for wildlife-2016. Proceedings RMRS-P-77; Albuquerque, NM; 2016 October 12-14. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 31-33. [95916]
250. Wiesenborn, William D. 2020. Phenology and description of a new species of Hoplothrips (Thysanoptera: Phlaeothripidae) in Mojave Desert leaf litter. The Pan-Pacific Entomologist. 96(1): 21-30. [95988]
251. Wiggins, Ira L. 1980. Flora of Baja California. Stanford, CA: Stanford University Press. 1025 p. [21993]
252. Wisdom, Charles S.; Gonzalez-Coloma, Azucena; Rundel, Philip W. 1987. Phytochemical constituents in a Sonoran Desert plant community. In: Provenza, Frederick D.; Flinders, Jerran T.; McArthur, E. Durant, compilers. Proceedings--symposium on plant-herbivore interactions; 1985 August 7-9; Snowbird, UT. Gen. Tech. Rep. INT-222. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: 84-87. [7401]
253. Wright, Michael E. 1973. Analysis of habitats of two woodrats in southern New Mexico. Journal of Mammalogy. 54(2): 529-535. [70345]
254. Young, James Albert; Young, Cheryl G. 1986. Collecting, processing and germinating seeds of wildland plants. In: CPR-4592. College Station, TX: Texas Agricultural Experiment Station. Portland, OR: Timber Press. 236 p. [12232]
255. Zouhar, Kris. 2003. Tamarix spp. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: https://www.fs.usda.gov/database/feis/plants/tree/tamspp/all.html. [96488]
256. Zouhar, Kris. 2005. Elaeagnus angustifolia. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: https://www.https://www.fs.usda.gov/database/feis/plants/tree/elaang/all.html. [96187]
257. Zouhar, Kristin; Smith, Jane Kapler; Sutherland, Steve; Brooks, Matthew L. 2008. Wildland fire in ecosystems: Fire and nonnative invasive plants. Gen. Tech. Rep. RMRS-GTR-42-vol. 6. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 355 p. [70897]

